Terminología descriptiva de las plantas
Los términos aquí citados corresponden a la terminología descriptiva de las plantas, que sirve para comprender las secciones de Descripción que aparecen en los artículos sobre taxones de plantas.
La terminología comienza a conocerse con un campo de estudio que trata sobre las estructuras normalmente visibles a ojo desnudo o con herramientas sencillas, como una lupa de mano, de los órganos de las plantas.[1] Los términos involucran las estructuras presentes en las plantas vasculares (helechos y angiospermas), y la terminología es muy extensa en lo que respecta a la flor de angiosperma, que es el grupo más diverso de plantas. Normalmente los briófitos (musgos y afines), que son plantas (son briofitas) pero con una morfología de origen evolutivo diferente, están descritos en un glosario aparte, como también tienen su propio glosario las estructuras relacionadas con la reproducción y otras más específicas de los helechos y de las gimnospermas. La morfología de alguna manera está integrada (se va diferenciando a medida que se disminuye la escala) con la anatomía[1] (o histología) de las plantas, que es el estudio de los tejidos y órganos a nivel celular, para lo cual se hace uso principalmente de preparados que se visualizan en un microscopio, para los que diseccionar el órgano idealmente embebido en un micrótomo, montarlo en un portaobjetos y teñirlo puede ser consumidor de tiempo (la definición de "anatomía" es diferente en plantas y animales).[2] La morfología forma la base de las descripciones taxonómicas y en general constituye el conjunto de datos más importante en la construcción de claves de identificación, ya que son los datos más rápidos y económicos de tomar y requieren de poca o ninguna técnica.[1]
Los términos citados aquí son en gran parte descendientes de aquellos usados por botánicos y herboristas en el pasado, empezando quizás por el antiguo griego Teofrasto, uno de los primeros en usar terminología técnica.[1] Muchos términos fueron heredados de la época del latín clásico o del griego antiguo latinizado, pero han evolucionado con los años: algunos fueron redefinidos, muchos fueron inventados —muchas veces en esos mismos idiomas—, y algunos descartados con el tiempo.[1] Especialmente desde la invención del microscopio se ha modificado la terminología en forma significativa.[1] El problema más grande que puede encontrar un usuario es que no hay consenso entre autores acerca de la definición precisa de muchos términos, por lo que, al utilizar una flora particular, debe ser chequeado el glosario de esa flora para corroborar el uso de los términos aquí aprendidos.[1] Los términos citados aquí son en gran medida derivados y clasificados, con algunas excepciones, de Radford et al. (1974[3]), véase también Bell (1991[4]), citados en el capítulo de morfología de Simpson (2005[1]) que es la base de este artículo (véase también las recomendaciones de Judd et al. 2007[5] en la sección de bibliografía).
La variabilidad es formalizada por los taxónomos en forma de caracteres "tipológicos"[6] que son puntos de anclaje del continuum de variación -una simplificación de la apreciación gestalt[7] del taxónomo que los utiliza, por lo que una descripción en palabras nunca será suficiente para abarcar la variabilidad del carácter en una especie o los detalles de cada carácter en la misma, de los que la única manera de aprenderlo es observando ejemplares de la misma especie; pero si la descripción dispone de un glosario con los límites de cada término morfológico claramente expresados, es importante repasarlo y extraer del texto toda la información que se haya racionalizado sobre el taxón, porque de los atributos que lo delimitan, los que son caracteres son una unidad evolutiva que puede tener una función o ser una restricción que explique una parte del plan corporal del taxón.
Una aproximación morfológica normalmente visualiza a la planta como estructurada en órganos altamente correlacionados con funciones específicas,[1] de forma que a partir de la función se establecen los límites del órgano,[1] y se dividen en secciones que describen la variabilidad encontrada en cada uno de ellos. Este artículo está dividido en tres secciones, la introducción a los órganos de las plantas, «Estructura de la planta» y «Terminología general». La sección de «Estructura de la planta» incluye caracteres y sus estados del carácter de órganos y partes de la planta específicos. Sigue una sección de «Terminología general» que lista caracteres y sus estados que pueden ser usados para describir una variedad de órganos de las plantas. La introducción a los órganos de las plantas fue ampliada solo en su sentido morfológico, al estilo de Bell (1991[4]) luego de la enumeración de los mismos, del resto, los agregados tomados de una fuente diferente de Simpson (2005[1]) son citados en línea, o en letra pequeña. Los agregados provienen principalmente de Judd et al. (2007[5]).
Introducción: los órganos de la planta
editar- Los artículos complementarios son Tracheophyta (las plantas vasculares) y sus derivados, como Pteridophyta (helechos y afines), Spermatophyta (las plantas con semilla) y Angiospermae (las "plantas con fruto", mal llamadas "plantas con flores").
Introducción a los órganos de la planta
editar.svg)
.svg)
En esta sección se introducen los órganos de las plantas y hasta qué nivel de detalle los describe el campo de la morfología.
Entre las plantas terrestres, la morfología del organismo multicelular, tanto de la generación gametofítica como de la generación esporofítica, divergió tempranamente en briofitos en sentido amplio (musgos y afines) y plantas vasculares (helechos y afines, gimnospermas y angiospermas); los briofitos normalmente tienen un glosario aparte y en este no serán tratados. De las plantas vasculares, en helechos y afines los gametófitos, si bien visibles a ojo desnudo, son poco importantes en su correlación morfología/taxonomía, y no serán tratados tampoco en este glosario, los de las gimnospermas y angiospermas son demasiado pequeños para ser tratados en morfología, y lo único que se observa de ellos son las estructuras del esporófito de la generación anterior que los encierran: el grano de polen (que encierra al gametófito masculino) y el óvulo (que encierra al gametófito femenino). El esporófito de estos grupos en cambio se desarrolla hasta ser conspicuo y nutricionalmente independiente del gametófito, y puede verse como una modificación del mismo esporófito ancestral "generalizado", que consta de sólo unos pocos órganos que a lo largo de la evolución fueron ricamente modificados en cada grupo.
Los órganos del esporófito al nivel más ancestral en plantas vasculares vivientes son la raíz[1] subterránea, el vástago[1] aéreo, y un sistema de haces vasculares que los vincula. El vástago a su vez está subdividido en tallo,[1] hojas[1] (megafilos o microfilos) (que a su vez contienen venas[1] continuas con el sistema vascular de la planta), más los órganos encargados de la reproducción, llamados esporangios.[1]
En plantas vasculares (exceptuando el grupo basal, las licofitas, que por practicidad también suelen tener su propio glosario), los esporangios siempre se encuentran en las hojas. La función del tallo es sostener y elevar hojas y esporangios, la de su sistema vascular, el transporte entre raíces y los órganos que eleva;[1] la función de las hojas es la fotosíntesis, lo que las vuelve también el sitio principal de intercambio gaseoso y transpiración; la función de la raíz es la absorción de agua y sales, y nitrógeno y fósforo asimilables en estados disponibles, disueltos en ella.[1] Raíz y tallo crecen a partir de un meristema apical (en su punta), que dejan los tejidos en formación detrás de ellos. Los extremos de raíz y vástago se especializan en proteger ese meristema, el órgano que protege el extremo de la raíz (la caliptra) es muy pequeño para ser visto en morfología, el que protege el extremo del vástago es la yema.[1] Los vástagos se ramifican en nuevos módulos, los módulos nuevos pueden especializarse, y reciben nombres nuevos de órganos. En helechos no (no hay ramificación), pero en espermatofitas los módulos o ramas portadoras de hojas fértiles (con esporangios) siempre están especializados, son ramas con las hojas dispuestas muy apretadamente a lo largo del tallo (de entrenudos cortos), es decir son braquiblastos, que se agotan después de dar la última hoja (de crecimiento determinado), que reciben el nombre de flor[1] (en castellano, la definición del inglés flower es diferente), a su vez la rama portadora de flores puede estar especializada en una inflorescencia.[1]
En espermatofitas, el esporangio femenino está encerrado por uno o dos tegumentos protectores en conjunto formando un óvulo[1] que después de la fecundación al madurar se vuelve una semilla, ya que la única espora y gametófito femenino formados en él nunca lo abandonan.[1] La semilla, o un órgano que la contiene, tiene la función de dispersión en gimnospermas. En angiospermas, las semillas al madurar se encuentran encerradas en el fruto, formado al mismo tiempo que ellas después de la fecundación a partir de tejidos que las rodean (principalmente los de la hoja fértil femenina o carpelo, que en este grupo las "envasan" como un tejido extra de protección y las ocultan de la herbivoría), y es el fruto, o una unidad que lo contiene o es parte de él, el que tiene la función de dispersión.[1]
En helechos no está dividida la sexualidad del gametófito, los gametófitos son todos bisexuales, por lo que no ocurre polinización, las gametas masculinas y femeninas se encuentran en el mismo gametófito; en espermatofitas, la polinización es el transporte del gametófito masculino hasta el óvulo o sus cercanías de manera de facilitar el encuentro entre los gametófitos y la fecundación. "Polinización" proviene de "polen", el grano de polen es el gametófito masculino encerrado en la espora que le dio origen, a la que todavía no ha abandonado.
En las angiospermas algunas hojas y partes de la flor se especializaron en órganos nuevos, que la adaptan a síndromes de polinización animales: los pétalos con función de atracción, los sépalos protectores, las partes de los carpelos: estigma (con función de recibir el polen y hacerlo germinar), estilo (con función de ser penetrado haustorialmente, alimentándose de él a medida que avanza, por el gametofito masculino) el ovario que contiene los óvulos, y partes de los estambres (la hoja fértil masculina): la antera (el conjunto de los esporangios masculinos, con la función de abrirse y exponer el polen al madurar), y normalmente un filamento que la eleva. Ésta es la lista de órganos como dada en Simpson (2005[1]) y traducida al castellano. Debido a que algunos grupos evolucionaron de forma más espectacular que otros, muchas veces cambiando drásticamente la forma y función de sus órganos, a veces es dificultoso diferenciar su origen sin un análisis detallado de los tejidos y la ontogenia y sin una comparación con grupos emparentados, muchas veces no podemos afirmar que un carácter que observamos en una planta es una "hoja" modificada si no nos lo indica un estudio previo.[8] En ese sentido es útil la distinción que hace la terminología entre las estructuras según su origen ancestral, y las estructuras que poseen esa función pero tienen otro origen del desarrollo (por ejemplo, la diferencia entre un "pétalo", que puede parecerlo o no, y un órgano "petaloideo" con función de pétalo, siendo virtualmente cualquier otra pieza capaz de evolucionar a petaloidea).
Las plantas vasculares vivientes, originadas en el ambiente terrestre, desarrollan un vástago aéreo y una raíz subterránea, los dos a partir de un meristema apical, un grupo de células indiferenciadas que se dividen por mitosis dejando los tejidos en formación detrás de ellas. Los dos meristemas están orientados a desarrollarse hacia arriba y hacia abajo respectivamente (poseen geotropismo negativo y positivo respectivamente). Esta especialización entre una parte aérea, que se desarrolla en busca del sol y el aire, y una parte subterránea, que se adentra en la tierra en busca de agua y sales, es lograda gracias a la comunicación entre ellas provista por los haces vasculares, los que le dan el nombre al grupo. Los haces vasculares son haces de apilamientos de células que recorren todo el largo de tallo y raíz, muchas veces están muertas a la madurez (cuando son funcionales), a través del interior de las cuales se transportan líquidos y solutos en solución (minerales, nutrientes como carbohidratos, hormonas...), y cuando es necesario los haces se ramifican de forma cada vez más fina hasta llegar a las inmediaciones de las células de todas las partes vivas de la planta, como se observa en la venación de las hojas. El xilema está formado por células muertas, de las que sólo quedan las paredes celulares formando un tubo que transporta el agua con sales minerales y otros solutos disueltos absorbida por las raíces (está asociado a células vivas que lo rellenan cuando se accidenta o envejece). El más delicado floema está formado por células vivas, que transportan la "savia elaborada" a través de su citoplasma, y se destruye después de envejecer y dejar de ser funcional. Las plantas, descendientes de algas que no contaban con ese impedimento, necesitan el intercambio gaseoso con el aire, ya que realizan fotosíntesis y respiración celular, pero son los momentos en que más agua pierden por transpiración. El agua transportada por el xilema desde las raíces se evapora al llegar a la superficie en contacto con el aire cercana a los extremos de los tubos de conducción. Una planta terrestre sin protección se desecaría en pocos minutos al Sol. Las plantas terrestres evitan la desecación en todas sus partes verdes gracias a que su epidermis, de una célula de espesor, es secretora de una capa cerosa que la impermeabiliza, la cutícula. La cutícula solo es interrumpida por los ostíolos (poros) delimitados por las células especializadas de los estomas, que dan a una cámara subestomática en la que el intercambio de gases ocurre sin la barrera de una cutícula gruesa. Los estomas están formados por dos células de la epidermis (células oclusivas) que cuando se modifican abren o cierran el ostíolo de forma acoplada con la situación metabólica de la planta en relación al ambiente. En el interior de tallo y raíz pueden haber quedado todo a su largo tejidos meristemáticos, o con el potencial de volver a serlo, llamados en conjunto meristemas laterales, responsables de un crecimiento posterior en grosor, llamado crecimiento secundario. Durante el crecimiento secundario, que puede presentarse en espermatofitas, la epidermis se rompe y es reemplazada por tejido formado debajo de ella, cuyas paredes en las capas celulares externas, en general muertas a la madurez, están impregnadas por compuestos complejos con una alta cantidad de ceras que lo impermeabilizan llamados suberina.
En el contexto de las descripciones morfológicas, por corteza se hace referencia al aspecto externo de las partes leñosas (de textura dura) de las plantas, en general coinciden con las protegidas por tejido de algún tipo de crecimiento secundario, sea este anómalo o no, pero también pueden volverse leñosas (de textura dura) las partes externas de los tallos por engrosamiento (lignificación, depósitos de lignina) de las paredes celulares al final de un crecimiento primario, como sucede en palmeras. El súber del crecimiento secundario respira por lenticelas, zonas con aspecto de verrugas[5] en la corteza en las que las células poco suberificadas presentan grandes espacios intercelulares que funcionalmente son poros, que permiten el intercambio gaseoso entre el exterior y las partes vivas internas que en esas partes de la planta es necesario principalmente para la respiración celular. Los meristemas laterales del crecimiento secundario también forman, en las capas más internas, nuevos haces vasculares de conducción que aumentan el tamaño de la copa y del sistema radical. Los haces vasculares de conducción de agua y sales (xilema primario, y después secundario), y las fibras a las que están asociados, son de por sí leñosos (endurecidos) y pueden formar una parte importante de la dureza del tallo interior a la corteza. Las fibras son células alargadas de paredes gruesas, paralelas al haz vascular, que crecen de forma intrusiva alcanzando hasta varios centímetros de largo, y son de hecho un componente principal de la fibra comercial. A medida que continúa el crecimiento secundario y los meristemas de los haces vasculares avanzan, formando una circunferencia de diámetro cada vez mayor, normalmente los tejidos más antiguos de conducción van dejando de ser funcionales y el xilema antiguo y relleno junto con sus tejidos asociados se van abandonando hacia el interior del tallo, quedando como tejidos de sostén y evidenciando su doble función. El más delicado floema conductor de azúcares se encuentra del lado externo de los meristemas, también está asociado a fibras, pero cuando envejece es comprimido por la presión que ejercen el xilema y el nuevo floema en formación más internos y por la resistencia que le oponen los tejidos corticales más externos, de manera que va dejando de ser funcional, y eventualmente se pierde junto con la corteza. Los meristemas laterales formadores de la corteza suberificada se regeneran en tejidos más internos, cercanos al floema funcional. Meristemas y haces vasculares, epidermis, estomas, el resto de los tejidos de la dermis, las diferentes formas de crecimiento secundario y engrosamientos del tallo, las texturas (leñosidad) y los tipos celulares de sostén se estudian a nivel celular en la anatomía vegetal,[2] también llamada histología vegetal en castellano. Las hormonas vegetales y los solutos y compuestos transportados por xilema y floema son parte de lo que se estudia en fisiología vegetal.
- El tallo
_foto_04.JPG)

_y_yemas_-_etiquetas.jpg)
El tallo se volvió postrado en las pteridofitas ancestrales, a lo largo del que se originan las hojas y las raíces adventicias. En los demás grupos (gimnospermas y angiospermas) el tallo es ancestralmente erguido.
Las ramas se forman en plantas vasculares ancestrales por división del meristema apical del tallo en dos (división dicotómica); en helechos y afines no se observa ramificación, y en espermatofitas, la ramificación es axilar, esto es que el meristema apical del vástago deja detrás de él zonas meristemáticas hacia el lado externo, justo debajo de la epidermis, observables al principio como abultamientos en el cilindro del tallo, llamadas primordios foliares —que se diferenciarán en hojas con los caracteres anatómicos de las eufilofitas— y los primordios de la yema en su axila (en el ángulo entre la hoja y el tallo).[9] Las hojas se desarrollan inmediatamente, las yemas pueden presentar un período de dormición. La sección del tallo en la que se inserta una o más hojas se llama nudo, y todo el largo del tallo comprendido entre dos nudos es un entrenudo. Las hojas programadas para tener una vida más corta que la rama que las porta, poseen en su base una zona de abscisión de forma que al caer la hoja deja una cicatriz en el tallo llamada cicatriz foliar cuyo aspecto puede ser parte de las descripciones morfológicas. La raíz primaria también se ramifica, pero no se ven primordios desde el exterior, ni posee los nudos con las yemas y hojas o cicatrices foliares que caracterizan al vástago. Los primordios de las raíces laterales se originan en tejidos interiores a la dermis y son visibles recién cuando rompen los tejidos de protección para abrirse paso a través de ellos. Los órganos que no se originan en los sectores mencionados se llaman adventicios. Son adventicias por ejemplo, las raíces que se originan en los nudos del tallo de las plantas rastreras, las estoloníferas y las rizomatosas.
- La hoja

_foto_03_planta_A.JPG)
.jpg)

Los licofilos de las licofitas (el grupo basal) se diferencian morfológicamente de los eufilos ("hojas verdaderas") del resto de las plantas vasculares por su tamaño y porque los inerva un único haz vascular, además de poseer diferencias anatómicas que aquí no serán expuestas; este glosario tratará sobre la diversidad de los eufilos, megafilos (de lámina amplia y venación ramificada) cuya lámina puede originarse de meristemas diferentes dentro de las eufilofitas. En algunos grupos se perdieron los megafilos secundariamente (después de haberlos adquirido), convergiendo con las licofitas en la morfología de un microfilo.
Normalmente los megafilos, de los que tratará este glosario, están compuestos por pecíolo y lámina o limbo, el pecíolo con la función de alejar la lámina del tallo y a veces torcerse y girar para orientarla de cara al sol; la lámina con la función de fotosíntesis, respiración y transpiración. Las hojas pueden ser envainadoras, de manera que las inervan más haces vasculares en su inserción al tallo. La vaina puede ser abierta o cerrada, cerrada es que sus extremos están fusionados del otro lado del tallo, pero una vaina puede rodear todo el tallo y ser abierta. Las hojas pueden presentar un par de apéndices a los lados de la base llamados estípulas, que pueden estar ricamente modificadas. La yema en su axila permite diferenciar una hoja simple de una compuesta en espermatofitas, los folíolos de las hojas compuestas pueden ser muy parecidos a las hojas simples, pueden hasta poseer peciolulos. Como el resto de las partes vivas de la planta, las hojas son inervadas por haces vasculares de transporte que en el caso de las hojas, por su grosor en relación con las mismas protruyen de la lámina o son observables a simple vista, por eso los patrones que forman entran dentro del campo de la morfología y son llamados venas o nervios.
La mayoría de las hojas son bifaciales:[5] tienen dos caras bien definidas y diferentes, normalmente una adaxial (ontogenéticamente ante el tallo) y una abaxial (ontogenéticamente alejada del tallo), que si la hoja no está resupinada (no gira sobre su eje) coinciden con la faz anterior y la posterior respectivamente, y en un tallo perpendicular al suelo (ortótropo), con la faz dorsal y la ventral de una hoja aplanada dorsoventralmente, en relación al cielo y al suelo ("ventral" es ante el suelo).
- Los esporangios
Los esporangios pueden ser muy variados en los glosarios de helechos donde su morfología y los patrones con los que se distribuyen en la hoja portadora son relativamente económicos de tomar y su uso enfatizado taxonómicamente,[9] pero no se tratarán en este glosario; las ramas especializadas en portar los esporangios de las gimnospermas, como los conos de las coníferas, poseen una morfología muy diferente de las de angiospermas y tampoco se tratarán en este glosario.
.svg)
_ovarios.svg)
_atardecer_anterior_a_la_antesis,_comienza_la_dehiscencia_y_se_observa_polen,_etiquetas.jpg)
_estigmas.svg)
_estilos.svg)
_-_6_-_9,57hs_hormigas,_acercamiento,_etiquetas.JPG)


Las angiospermas comparten con las gimnospermas que poseen estambres y carpelos, las hojas fértiles, portadoras de los esporangios masculinos y femeninos respectivamente (en castellano), y en ambos grupos están tan modificados que normalmente no son reconocibles como hojas sin el conocimiento previo; ambos se presentan en ramas de entrenudos cortos de crecimiento determinado llamadas flor (en castellano), las que se especializan en algún "síndrome floral" (adaptación a una estrategia particular de reproducción). En espermatofitas, por "reproducción" se implica polinización (transporte por agentes externos del grano de polen, que porta al gametofito masculino, hasta las inmediaciones del óvulo que porta al gametofito femenino), el encuentro entre los dos gametofitos, fecundación y formación del embrión nutrido por el gametofito femenino y su esporófito madre hasta la formación de la semilla madura (lista para germinar), por lo que en angiospermas, en el síndrome floral también se encuentra la morfología del fruto al comenzar su formación, se puede decir que el ovario y las partes accesorias que formarán el fruto son un "fruto inmaduro" ya justo después de la fecundación, que al madurar está listo para dispersión, o lo está una unidad que lo contiene o es parte de él.
En la flor de angiosperma, las hojas del síndrome floral o "piezas" se disponen alrededor del tallo especializado en este orden: los sépalos (piezas protectoras, verdes) que en conjunto forman el cáliz, los pétalos (piezas llamativas de atracción de polinizadores animales) que en conjunto forman la corola (ambos grupos en conjunto son el perianto), los estambres que en conjunto forman el androceo, y los carpelos que en conjunto forman el gineceo, este a su vez conformado por una o varias unidades funcionales llamadas pistilos —cada pistilo formado por uno o más carpelos—. La región del tallo en la que están insertas las piezas es llamada receptáculo, o eje floral o tálamo. El receptáculo puede ser una continuación del tallo en su morfología o puede ser expandido, puede tener forma de plato o cabezuela, o puede volverse una concavidad, como en las flores de ovario ínfero, etc. El orden en que se disponen las piezas en el eje nunca varía —alguna puede faltar—, y el eje floral nunca deja de ser determinado (nunca deja de agotarse después de dar la última pieza), pero es común que varíe la forma y función de las piezas florales y que su morfología sea una parte importante de las descripciones taxonómicas. La forma del receptáculo, la posición y número de las piezas florales en él y cómo se relacionan entre ellas (las diferencias relativas en tamaño, forma, si hay piezas florales fusionadas o faltantes) forman síndromes florales "generalizados" que suelen ser parte de las descripciones taxonómicas de grupos que se ubican en la categoría de familia,[cita 1] otra forma de decirlo es que típicamente definen familias de plantas (véase fórmula floral y diagrama floral).
En el estambre, en el interior de cada esporangio masculino, se forman granos de polen, es la espora masculina cubierta de material proveniente del esporófito, en el interior de la cual se desarrolla el gametofito masculino hasta el momento en que el grano de polen ya está "maduro", listo para ser liberado —el gametofito masculino es "endospórico"— (Simpson 2005[1] lo llama gametofito inmaduro, porque en el momento de su liberación aún no fue diferenciado). En el carpelo a su vez se forma el o los óvulos (en la figura en negro), un esporangio femenino rodeado de un (gimnospermas) o dos (angiospermas) tegumentos protectores provenientes del esporófito, que en su interior forma una única espora femenina, la que a su vez en su interior forma un único gametofito femenino -también endospórico. Los nombres de género "masculino" y "femenino" fueron asignados debido a que la gameta femenina nunca abandona el gametofito femenino, y es la que está rodeada de tejidos nutricios. En cambio las células espermáticas nadan hasta la gameta femenina en grupos primitivos, y también en espermatofitas son acercadas hasta las proximidades de la gameta femenina primero por agentes externos que transportan el gametofito masculino (proceso llamado polinización), y en angiospermas el gametofito masculino se termina de acercar activamente por crecimiento haustorial (adentrándose y alimentándose de los tejidos que atraviesa) en el carpelo hasta el gametofito femenino y su gameta.
En angiospermas, el carpelo está plegado encerrando el óvulo dentro de él y ofrece en su lado externo una superficie especializada en recibir el polen llamada estigma o superficie estigmática, el polen que se deposita en el estigma "germina", esto es, el gametofito masculino emite un "tubo polínico" —se desarrolla por elongación de sus células hacia afuera de la pared del polen penetrando por el estigma en los tejidos del estilo macizo (no hueco) que es atravesado haustorialmente (alimentándose de él a medida que avanza) hasta el ovario y uno de sus óvulos, en el que penetra y encuentra el gametofito femenino—. Al emitir dentro de él un juego de cromosomas ocurre la fecundación, que en angiospermas es "doble", debido a que también debe fecundar —agregar un juego de cromosomas— al tejido del gametofito femenino que se desarrollará en un tejido de reserva que proporcionará los nutrientes al nuevo esporófito cuando se desarrolle: el endosperma. También los restos del esporangio pueden desarrollarse formando otro tejido de reserva: el perisperma. El mismo gametofito femenino es un tejido de reserva, relativamente significativo en gimnospermas. Formación de granos de polen y óvulos, el recorrido del tubo polínico y lo que ocurre después de la fecundación hasta la formación de la semilla madura (lista para la dispersión) son tratados fundamentalmente al microscopio en el campo de la embriología de las plantas,[11] y en particular el polen maduro (liberado para la polinización), como se presenta en forma ubicua en la Tierra y sus capas externas persistentes son muy ornamentadas, dan información taxonómica (que puede separar grupos claramente) y se mantienen durante muchos años sin degradarse, ofrece mucha información utilizada en campos como filogenia, paleontología, biogeografía, identificación (determinación) de especies y hasta biología forense; el campo que describe el polen de cada especie es llamado palinología.[12]
Una rama que porta más de una flor es una inflorescencia, el tallo en su origen es el pedúnculo (no confundir con el pedicelo de cada flor individual). El "síndrome floral" adaptado a una estrategia particular de polinización no siempre involucra a una flor individual, a veces puede ser una inflorescencia que funciona como una única flor, como en las flores compuestas (llamadas así desde antiguo porque son "flores compuestas por flores") o asteráceas, como el girasol, la margarita y el panadero; en los que cada "semilla" deriva de una única flor, o en las aráceas, como la cala, en la que el "pétalo" de la cala es en realidad una bráctea que da inicio a la inflorescencia protegida por ella. A veces el síndrome floral es sólo un sector radial de una flor simétrica, como en el lirio (Iris), donde una única flor da lugar a 3 construcciones florales bilabiadas" (con "dos labios", dos piezas petaloideas, una dorsal y una ventral) que encierran un estambre y un estigma cada una, pareciendo toda la flor una inflorescencia.
- La semilla

En helechos y afines el nuevo esporófito se desarrolla directamente sobre el gametofito en el que ocurrió la fecundación, al principio nutrido por este —es un embrión— pero enseguida se independiza, sin entrar en dormición. En las "plantas con semilla" (gimnospermas y angiospermas) una vez formado el cigoto en el carpelo, el nuevo esporófito al desarrollarse protegido y alimentado por el gametofito que lo originó es correctamente llamado embrión, los tegumentos del óvulo provenientes de su esporófito madre maduran en tegumentos de protección en la semilla, y pueden madurar también los tejidos de reserva —endosperma y perisperma—; hasta que en un momento se detiene su desarrollo y el nuevo esporófito entra en latencia dentro de sus tegumentos y tejidos de reserva: es la semilla madura, lista para la dispersión. En morfología solo se detallan algunos caracteres del óvulo y la semilla, en general los visibles sin ayuda del microscopio —por ejemplo, la ubicación y forma del hilo, que es la cicatriz que dejó al caer el funículo, el cordón que unía la semilla a la placenta en la pared del carpelo—, y el aspecto a ojo desnudo del embrión al abrir los tegumentos, en el que ya se pueden apreciar morfológicamente el primer vástago y la primera raíz, el primero con hojas llamadas cotiledones, típicamente engrosados y reservantes. La semilla es el órgano de dispersión y colonización de nuevos ambientes en gimnospermas, en angiospermas es el fruto.
- El fruto

.png)
En angiospermas el fruto (el "carpio") se desarrolla después de la fecundación a partir de un ovario. Pueden sumarse partes de las cercanías en la misma flor llamadas "partes accesorias", como el receptáculo en frutillas y manzanas.
El pericarpio es la pared del fruto maduro (listo para dispersión), es un término de la morfología del fruto, no de los tejidos de los que proviene. Por lo que el pericarpio podría incluir partes accesorias si las hubiera. Tradicionalmente se identifican en él 3 capas con funciones diferentes: el exocarpio externo de protección (la "cáscara"), el mesocarpio y el endocarpio interno de protección de la semilla. No siempre se diferencian las 3 capas.
Nótese que el fruto no necesariamente se corresponde con todo el gineceo de una flor individual, los frutos podrían derivar de cada pistilo individual de una flor de pistilos libres.
Nótese también que "fruto" no es la "unidad de dispersión" como en el lenguaje coloquial. La unidad de dispersión podría ser un polifruto, un fruto o polifruto con partes accesorias provenientes de partes externas a la flor (como en Juglans regia, la nuez en lenguaje coloquial), o una infrutescencia, como en el ananá (Ananas comosus), cuyo tallo se continúa después de dar la infrutescencia en un "penacho" (una roseta) de hojas fotosintéticas. Las unidades de dispersión que mejor conocemos son las que están adaptadas a la estrategia ecológica de recompensar con alimento a los mamíferos que en consecuencia transportan y depositan las semillas en sitios alejados de la planta original.
El síndrome floral que permite la reproducción de la planta (polinización, fecundación y dispersión) es parte de lo que se estudia en la biología reproductiva de las plantas[13] (Judd et al. 2007[5] en cambio menciona el campo más estricto de la "biología de la polinización" o biología floral).
- La plántula
La semilla dispersada puede necesitar de condiciones particulares que pueden ser que un animal la quite de su fruto, la escarificación (degradación de los tejidos del exocarpio) por el paso por el sistema digestivo de un animal herbívoro, o un período de frío, etc. antes de germinar. La germinación normalmente comienza con la absorción de agua. Los cotiledones juegan un rol crucial en este período, pueden mantenerse dentro de los tegumentos y funcionar como órgano de absorción del endosperma y otros tejidos nutritivos durante la germinación, o pueden volverse epígeos (ser elongados por sobre tierra) y fotosintéticos, o ser ellos mismos reservantes de nutrientes, o combinaciones de estas funciones. Desde la germinación hasta que termina de absorber las reservas que trajo consigo en la semilla y se marchitan sus hojas cotiledonares, el esporófito se encuentra en el estadio de "plántula". Si al finalizar este estadio la planta ya es independiente, seguirá desarrollándose hasta madurar (portar hojas fértiles) y reiniciar el ciclo: ya es una planta establecida.
La planta establecida es una planta juvenil, si llega a desarrollarse hasta tener la posibilidad de portar hojas fértiles (aunque todavía no lo haya hecho) es una planta adulta. Las plantas pueden ser adultas y no madurar nunca, como las que se crían fuera de su nicho ecológico en ambientes donde nunca encontrarán las condiciones para portar hojas fértiles. Las plantas en condiciones diferentes también pueden mantenerse juveniles por el resto de su vida, como los potus de interior, que nunca llegan a desarrollar los caracteres de la especie adulta. La planta madura es la que fructificó y dio unidades de dispersión viables al menos una vez.
- La sucesión de hojas
.png)
Las hojas primordiales son las primeras hojas que nacen por encima de los cotiledones de la planta joven, en angiospermas tienen una forma diferente de las hojas de la planta adulta, son más pequeñas y en general más simples (por ejemplo en plantas de hojas compuestas, tienen menor número de folíolos que las hojas que aparecerán después). Una vez que el tallo aumentó de grosor y cuenta con el número de haces vasculares como para poder inervarlas, y por todo el resto de la vida de la planta, aparecen las hojas vegetativas o nomófilos, las hojas fotosintéticas de la planta adulta, características de cada especie.
La sucesión a hojas más grandes y complejas también puede darse en las ramas individuales a medida que aumentan de grosor.
Los profilos son las hojas del primer nudo sobre un eje lateral (más estrictamente un eje axilar, originado en la axila de una hoja, ya que a veces un eje lateral es el mismo tallo que se geniculó -se torció-, como en los ejes simpodiales), no tienen yema en su axila, son dos en dicotiledóneas y uno en monocotiledóneas, y también son diferentes de los nomófilos, normalmente más pequeños (muchos autores los llaman "bracteolas"). Sobre el eje axilar después de los profilos pueden desarrollarse nomófilos con la respectiva yema en su axila u otras hojas; si aparecen los antofilos (las piezas florales), entonces es un "eje floral". Cuando el mismo eje pasa del estado vegetativo al estado floral, a menudo el cambio es anunciado por una modificación en la forma de las hojas. Por ejemplo el limbo puede reducirse, la hoja volverse sésil (sin pecíolo), y la coloración puede cambiar. La palabra general para las hojas que no son nomófilos es brácteas, muchas brácteas no tienen yema en su axila.[14]
Estructura de la planta
editarHábito, duración, hábitat y distribución
editarHábito
editarEl hábito o hábito de crecimiento, o a veces porte de la planta se refiere a la forma general de la planta, lo cual abarca una serie de componentes como la duración del tallo (si es estacional, perenne), el patrón de ramificación, el desarrollo, la textura —si alguna de sus partes está endurecida, leñosa—, mediante qué estrategia sobrevive la época desfavorable, y cuál es su estrategia de establecimiento. La mayoría de las plantas puede ser claramente designada como hierba, guiadora, arbusto o árbol (con algunas subcategorías), sin embargo, algunas especies son difíciles de acomodar en alguna categoría.
Algunos tipos de hábito de la planta y tallo son los siguientes:

_-_%22Lavanda%22_probably_Lavandula.JPG)

- Una hierba (planta herbácea) es una planta en la cual todos los vástagos sobre la superficie de la tierra, sean vegetativos o reproductivos, se mueren al final de la estación de crecimiento anual, y nunca llegan a ser leñosos. Si bien los vástagos sobre la superficie son anuales, la hierba en sí misma puede ser anual, bienal (que vive por dos años), o perenne. Las hierbas que sobreviven por más de una estación de crecimiento poseen órganos subterráneos reservantes, cuando es así persisten mediante yemas que permanecen debajo de la superficie del suelo y son llamadas geófitas. Puede también haber hierbas siempreverdes.
- Se llama cespitosa la hierba que tiene múltiples tallos aéreos y muy cortos, originados en la base (cerca del suelo), las plantas cespitosas poseen un aspecto de cojín muy ramificado. Son comunes los pastos cespitosos (familia Poaceae). Los pastos cespitosos se ramifican muy densamente por macollos (ramificaciones de los pastos, con el primer nudo de diámetro muy pequeño) intravaginales (que se mantienen protegidos dentro de la vaina de la hoja en cuya axila se originaron), a diferencia de los que ramifican extravaginalmente (escapándose de la vaina) y son aptos para cobertura del suelo.[4]:182
- Un arbusto (planta arbustiva o fruticosa, frútice, frutescente, tallo frutescente) es una planta perenne, leñosa con muchos tallos principales que se elevan sobre el nivel del suelo, y ninguno especialmente engrosado.
- Un árbol (planta arborescente o arbórea, tallo arborescente) es una planta perenne, leñosa, con un tallo principal (el tronco) que se eleva sobre el nivel del suelo, el tronco puede ser ramificado o no. Si no se ramifica, o si se ramifica solo para dar una inflorescencia como en el caso de las palmeras cuyas inflorescencias son axilares (originadas en la axila de una hoja), se llama estípite.[15] En general el árbol es más alto que el arbusto. No todos utilizan estas definiciones de arbusto y árbol, por ejemplo algunos ecólogos diferencian entre arbusto y árbol solo por su altura, entonces es útil diferenciar el hábito de la planta y el del tallo.
- Un subarbusto (sufrútice, sufrutescente, planta sufruticosa, tallo sufrutescente), es un arbusto corto que es leñoso solo en la base, los primeros centímetros por encima del suelo, siendo el resto de la planta compuesta por ramas herbáceas, la planta nunca demasiado alta.
.jpg)
- Una planta acaulescente (un tallo de hábito acaulescente), a diferencia de las caulescentes, no poseen tallo ("caule") sobre el nivel del suelo más que el de la inflorescencia en el momento de la floración. Pueden poseer hojas fotosintéticas pero solo al ras del suelo, usualmente en una roseta basal, cuando es así son plantas en roseta. Las plantas acaulescentes son muchas veces hierbas bienales, en las cuales una raíz de almacenamiento se desarrolla en el primer año y la floración ocurre en el segundo; o hierbas perennes, en las cuales el tallo persistente en el que se insertan las hojas fotosintéticas se mantiene bajo tierra y protegido durante las condiciones ambientales desfavorables. El pedúnculo de la inflorescencia de una planta acaulescente se llama escapo, el escapo puede tener hojas, pero no bien desarrolladas, no son nomofilos, la planta en este caso se llama "escaposa".
- Escapos
-
 Escapo, señalado con flecha. En amarillo claro el tallo (escapo), en rojo y amarillo fuerte la inflorescencia, en verde las hojas, en marrón las raíces.
Escapo, señalado con flecha. En amarillo claro el tallo (escapo), en rojo y amarillo fuerte la inflorescencia, en verde las hojas, en marrón las raíces. -
 Escapos en Allium tuberosum.
Escapos en Allium tuberosum.


- Una planta de guía o guiadora es una planta con tallos elongados y débiles, con la estrategia ecológica de buscar el Sol lejos de su sitio de germinación sin necesidad de sostenerse por sí misma. En general se encaraman a un sustrato (son trepadoras) y se fijan mediante órganos de fijación (son enredaderas), pero no necesariamente son una ni la otra. También hay trepadoras no guiadoras, como en los ejemplos de hábitos vistos más adelante. Ver la terminología de la planta y sus órganos de fijación (guiadora herbácea, leñosa, liana, escandente, trepadora, guiadora rastrera, órganos de fijación: enredadera, voluble, zarcillo caulinar, zarcillo foliar, raíces adventicias de tipo pad adhesivo) en Planta de guía.

- Las plantas cuyas partes aéreas se apoyan en el sustrato cuando adultas se llaman postradas o de "tallo postrado", y se asume que si se las llama postradas no se está refiriendo a plantas guiadoras ni rastreras.[1] Si se apoyan en el sustrato pero el ápice se eleva verticalmente, se llaman decumbentes o de tallo decumbente.[1] Una planta guiadora también puede tener tallo decumbente. Si no son postradas ni decumbentes, son erguidas (no confundir con tallo ortótropo y tallo plagiótropo, que significan perpendicular al suelo y paralelo al suelo respectivamente). Son plantas rastreras las que enraízan en los nudos en contacto con suelo húmedo. Son pastos rastreros la gramilla y el gramón (no confundir con los pastos rizomatosos, los rizomas se ven similares pero están bajo tierra y son reservantes, son pastos tipo césped). Las guiadoras también pueden ser además rastreras (enraizar en los nudos). Los estolones son un tallo al ras del suelo y alargado de propagación vegetativa, una planta estolonífera no se llama por eso postrada ni rastrera, son plantas (erguidas o no) con estolones. La nueva plántula al final de un estolón eventualmente se independiza de la planta madre, como en las "cintas" y en Fragaria (la frutilla o fresa/fresón). Como los estolones pueden crecer bajo tierra, a veces son llamados tallos subterráneos y parecen rizomas delgados y elongados, pero no son reservantes, y la planta terminal termina separada de la planta principal. Esto también los diferencia de las plantas rastreras, aunque hay una gradación entre estoloníferas y rastreras.
- Estoloníferas y rastreras
-
 Estolón. En amarillo claro el tallo, en verde las hojas. En marrón oscuro las raíces.
Estolón. En amarillo claro el tallo, en verde las hojas. En marrón oscuro las raíces. -
 Estolones en las frutillas o fresas.
Estolones en las frutillas o fresas. -
 Estolones en las "cintas", Chlorophytum comosum, que suspenden a las plantas hijas.
Estolones en las "cintas", Chlorophytum comosum, que suspenden a las plantas hijas. -
 Comparar con los "gramones" y "gramillas", según otros autores ya plantas rastreras.
Comparar con los "gramones" y "gramillas", según otros autores ya plantas rastreras.




Las plantas pueden tener estos hábitos por sólo una parte de sus vidas, como las hemiepífitas y las semitrepadoras:

- Las de hábito semitrepador[16][17][18] (por hemi-) o apoyante[16][18][19] inician su vida en el sotobosque de forma erguida y leñosa y parasitan mecánicamente los árboles solo al llegar al dosel, en donde se apoyan y entrelazan sus ramas, hojas, o inflorescencias[16] muchas veces ayudados por espinas o ganchos. Son arbustos apoyantes las rosas (Rosa),[17] probablemente Monstera y Philodendron (dos aráceas terrestres semileñosas), las arecáceas llamadas ratanes, y quizás el árbol del viajero (Ravenala madagascariensis).
- Las plantas estranguladoras o hemiepífitas leñosas,[17] a diferencia de otras plantas leñosas comienzan su vida como epífitas (germinan sobre otras plantas y crecen sobre ellas sin parasitarlas), y desarrollan raíces aéreas leñosas dirigidas hacia el suelo. Una vez las raíces llegan al suelo enraízan y se vuelven de sostén (la planta pasa a ser terrestre), con el tiempo las raíces alrededor del tronco de la planta hospedadora se engrosan y el hospedador muchas veces muere por competencia por la luz solar o quizás estrangulamiento al no tener cómo continuar su crecimiento secundario. Este hábito al madurar muchas veces les da un aspecto arborescente cuyo "tronco" es un sistema tallos-raíces estranguladoras.[20] Es encontrado en algunas Moraceae, en particular del género Ficus (las especies conocidas como "higueras estranguladoras").
- Plantas estranguladoras (hemiepífitas leñosas)
-
![Un estrangulador maduro puede tener un hábito arborescente, cuyo "tronco" está formado por un sistema raíces-tallos que se anastomosan en un proceso de injerto natural.[20]](//upload.wikimedia.org/wikipedia/commons/thumb/8/8f/Macau_Mt_Fortress_Museum_of_Macau_n_Banyantree.JPG/213px-Macau_Mt_Fortress_Museum_of_Macau_n_Banyantree.JPG) Un estrangulador maduro puede tener un hábito arborescente, cuyo "tronco" está formado por un sistema raíces-tallos que se anastomosan en un proceso de injerto natural.[20]
Un estrangulador maduro puede tener un hábito arborescente, cuyo "tronco" está formado por un sistema raíces-tallos que se anastomosan en un proceso de injerto natural.[20] -
 Las plantas estranguladoras comenzaron su vida como epífitas de raíces leñosas.
Las plantas estranguladoras comenzaron su vida como epífitas de raíces leñosas.
![Un estrangulador maduro puede tener un hábito arborescente, cuyo "tronco" está formado por un sistema raíces-tallos que se anastomosan en un proceso de injerto natural.[20]](/wiki/Archivo:Macau_Mt_Fortress_Museum_of_Macau_n_Banyantree.JPG)

- Las plantas carnívoras (en ambientes oligotróficos u oligótrofos, pobres en nutrientes, especialmente nitrógeno asimilable), están dotadas de dispositivos especiales mediante los cuales capturan y retienen pequeños animalitos, sobre todo insectos, los digieren parcialmente y los utilizan como fuente suplementaria de nitrógeno orgánico.[21] Las plantas carnívoras, junto con las parásitas son plantas total o parcialmente heterótrofas.[21] Tres tipos de hojas son adaptaciones muy especializadas de las plantas carnívoras. Las hojas "pitcher" están adaptadas como un contenedor, que contiene un fluido en su interior y funciona en la captura y digestión de animales pequeños. Taxones con hojas así son Darlingtonia, Nepenthes y Sarracenia. Las hojas tentaculares llevan numerosos pelos (tricomas) glandulares pegajosos que funcionan en la captura y digestión de animales pequeños, como en Drosera. Las hojas trampa ("trap") son las que mecánicamente se mueven después de ser disparadas por algún mecanismo, capturando en el proceso a animales pequeños para luego digerirlos, como en Dionaea muscipula.
- Hojas de plantas carnívoras
-
 Hoja "pitcher" en Sarracenia.
Hoja "pitcher" en Sarracenia. -
 Hoja tentacular de Drosera atrapando un insecto.
Hoja tentacular de Drosera atrapando un insecto. -
 Hoja trampa en Dionaea.
Hoja trampa en Dionaea.



- Las parásitas se desarrollan sobre otra planta, parasitándola.[21] La función de las raíces desaparece en muchos casos, habiendo éstas evolucionado a raíces haustoriales (haustorial: que crecen intrusivamente en el hospedador alimentándose de sus tejidos a medida que avanzan) que se conectan con el sistema vascular del hospedador. Las holoparásitas no son verdes ni casi fotosintetizan, carecen casi completamente de clorofila, presentan una reducción más o menos importante de los órganos vegetativos, las hojas se reducen a escamas amarillentas. Las hemiparásitas (por semi-) son plantas verdes fotosintetizantes que además poseen raíces haustoriales.[21]
- Las parásitas epífitas se fijan sobre el vástago del hospedante como Cuscuta[21] y Pilostyles[21] (no confundir con plantas epífitas, sin más adjetivos, son las que crecen sobre otras plantas sin parasitarlas).
- Las parásitas epirrizas se fijan sobre las raíces del hospedante, como la "flor de piedra", Lophophytum leandri, parásita de Piptadenia.[21]
- Plantas parásitas
-
 Cuscuta (amarilla, sobre el árbol), sus raíces haustoriales penetran al hospedador haustorialmente (alimentándose de él a medida que avanzan) hasta conectarse a su sistema vascular. Los estrictos controles de "libre de cuscuta" son necesarios para el transporte de vegetales en algunos países.
Cuscuta (amarilla, sobre el árbol), sus raíces haustoriales penetran al hospedador haustorialmente (alimentándose de él a medida que avanzan) hasta conectarse a su sistema vascular. Los estrictos controles de "libre de cuscuta" son necesarios para el transporte de vegetales en algunos países. -
 Viscum album, el muérdago, una hemiparásita (por semi-) parasitando un álamo y fotosintetizando.
Viscum album, el muérdago, una hemiparásita (por semi-) parasitando un álamo y fotosintetizando.


- Las plantas epífitas crecen sobre otra planta solo usándola de sustrato, sin parasitarla. Desarrollan caracteres de ambientes secos (son xeromórficas) debido a la velocidad a la que se escurre el agua de lluvia por ellas. Pueden tener raíces aéreas, son raíces adventicias que en general no llegan al suelo, y acumulan agua y minerales del aire o que se escurren sobre ellas (una definición muy amplia de raíces aéreas es la definición literal, toda raíz o porción de ella que está sobre el nivel del suelo[5]). Muchas raíces aéreas son verdes, fotosintéticas. Su ocurrencia se suele mencionar en los taxones Orchidaceae y Araceae, y también se encuentran en algunas Cyperaceae y Velloziaceae.[22] La epidermis de las raíces aéreas es una adaptación a la exposición al aire llamada velamen.[23] Este tejido, como las raíces aéreas, se originó varias veces independientemente en la evolución, es una expansión de la epidermis de muchas células de espesor, muertas a la madurez, que forman un tejido esponjoso que absorbe el agua y sobrelleva los períodos de sequía sin que se reseque el interior funcional de la raíz. Presenta un color blancuzco perlado cuando se reseca, y se transparenta al humedecerse mostrando el interior de la raíz muchas veces fotosintético. El velamen también puede presentarse en algunas orquídeas no epífitas emparentadas con éstas.
- Epífitas
-
 Una bromeliácea epífita ocupando de forma oportunista un cableado. Las bromeliáceas acumulan agua de lluvia en las bases envainadoras de sus hojas en roseta, que además son suculentas.
Una bromeliácea epífita ocupando de forma oportunista un cableado. Las bromeliáceas acumulan agua de lluvia en las bases envainadoras de sus hojas en roseta, que además son suculentas. -
 Velamen en una orquídea.
Velamen en una orquídea.



- Las plantas poikilohídricas o reviviscentes (a diferencia de las homeohídricas cuya hidratación es independiente de la humedad del ambiente) dependen de la humedad ambiente para mantenerse hidratadas. Pueden sufrir una desecación extrema, y cuando vuelve a estar disponible el agua se rehidratan y reanudan sus funciones normales. Entre ellas están las briofitas y algunas pteridofitas como Selaginella lepidophylla, la "planta de la resurrección" (Ingrouille 1992[24] citado en[25]) y Polypodium squalidum, especie nativa de Corrientes, Argentina.[25]
- Otras partes de la planta utilizadas en la descripción del hábito
La descripción del hábito puede incluir la de los estadios del desarrollo juveniles de la planta. La sucesión foliar desde las primeras hojas cotiledonares pasando por las hojas primordiales a las hojas "normales" vegetativas o nomofilos puede tener diferente morfología. La planta puede tener además otros tipos de hojas: puede tener heterofilia (dos morfos de nomofilos, por ejemplo por debajo y por encima del agua), hojas no nomófilos, como las protectoras de las yemas en dormición en la época desfavorable, pueden tener domatia ("casitas" para habitáculo de alguna especie simbionte de insecto), etc. Se describen más adelante en hoja.
La planta adulta también puede estar armada de espinas o aguijones. La terminología ("espina caulinar", "espina foliar", "aguijón") en su propio artículo, Espinas y aguijones (Botánica).
Una planta adulta puede tener partes reservantes, o puede tener partes suculentas, como los cactus del Nuevo Mundo y las euforbiáceas suculentas de África (tallo suculento)[1] o las crasuláceas de hojas suculentas. Las partes reservantes tienen un rol en la longevidad de la planta en hábitats con épocas desfavorables (ver en Duración). Se describen en su propio artículo, Órganos reservantes en las plantas.
- Un cladodio o filóclado es un tallo fotosintético, puede parecerse a una hoja, como en Opuntia (la tuna), Asparagus (los espárragos), y Ruscus. Los cladodios cumplen la función fotosintética que normalmente tienen las hojas, y pueden ayudar a disminuir la pérdida de agua.
- Cladodios o filóclados
-
 Cladodios. En amarillo claro el tallo, en verde las hojas. En rojo las flores.
Cladodios. En amarillo claro el tallo, en verde las hojas. En rojo las flores. -
 Cladodios en Opuntia.
Cladodios en Opuntia. -
 Los cladodios de Ruscus parecen hojas. Aquí con flores.
Los cladodios de Ruscus parecen hojas. Aquí con flores. -
 Las "ramas plumosas" de los espárragos son cladodios fotosintéticos, las hojas son pequeñas y escamosas.
Las "ramas plumosas" de los espárragos son cladodios fotosintéticos, las hojas son pequeñas y escamosas.




- Un braquiblasto ("rama corta"), o fascículo (en inglés "fascicle", "short shoot", "spur shoot", "dwarf shoot") es un tallo con entrenudos muy cortos. Los braquiblastos permiten la producción de hojas u otros órganos relativamente rápido, con mínimo tejido del tallo formado. Los braquiblastos pueden ser encontrados en plantas deciduas de climas de sequía, que producen un rápido crecimiento de hojas desde los braquiblastos después de la lluvia. Los braquiblastos crecen desde las yemas de tallos más típicos con entrenudos más largos, los últimos llamados, en contraste, macroblastos ("ramas largas", en inglés "long shoots").
- Braquiblastos
-
 Fascículo o braquiblasto. En marrón el macroblasto, en marrón claro la escama, en amarillo el braquiblasto, en verde las hojas.
Fascículo o braquiblasto. En marrón el macroblasto, en marrón claro la escama, en amarillo el braquiblasto, en verde las hojas. -
 Braquiblastos de Pinus canariensis y un macroblasto.
Braquiblastos de Pinus canariensis y un macroblasto. -
 Braquiblasto de Ginkgo biloba originado en un macroblasto.
Braquiblasto de Ginkgo biloba originado en un macroblasto.


- Raíces fúlcreas, raíces zancudas o zancos (en inglés prop roots o stilt roots), son adventicias, se originan sobre la superficie de la tierra, en general cerca de la base del tallo, penetran en la tierra verticalmente y se vuelven leñosas. En los casos más típicos se parecen a patas de araña, como en el pandano (Pandanus) y el maíz (Zea mays). Otro ejemplo son los manglares (Rhizophora).[20] Algunos autores dan ejemplos de estas raíces que no necesariamente se originan en la base del tallo sino en partes alejadas del origen de la planta y sobre las ramas, no se parecen a patas de araña, pero penetran verticalmente en el suelo con función de sostén,[20][26] como en Ficus benghalensis,[20] la "higuera de la India"[26].
- Raíces fúlcreas
-
 Raíces fúlcreas en el pandano, Pandanus.
Raíces fúlcreas en el pandano, Pandanus. -
 Raíces fúlcreas en el maíz (Zea mays).
Raíces fúlcreas en el maíz (Zea mays). -
 Las espectaculares raíces fúlcreas del manglar rojo le dan su apariencia característica, a veces se las encuentra como raíces cable.
Las espectaculares raíces fúlcreas del manglar rojo le dan su apariencia característica, a veces se las encuentra como raíces cable. -
 Las raíces aéreas alejadas del origen de la planta de algunas especies se insertan en el suelo como raíces fúlcreas.
Las raíces aéreas alejadas del origen de la planta de algunas especies se insertan en el suelo como raíces fúlcreas.

.jpg)


- Raíces tabulares[26] o tablares[27] (en inglés buttress roots o tabular roots) que pueden verse en algunos árboles, son raíces leñosas, engrosadas, que se originan en la base del tronco y están un poco enterradas pero sobresalen por sobre la superficie del suelo, extendiéndose horizontalmente en él hasta desaparecer completamente debajo de la tierra, y usualmente (pero no necesariamente) son verticalmente engrosadas (pareciendo tablas). Las "tablas" formadas por estas raíces reciben diferentes nombres: aletones, gambas, bambas.[27] Son raras en árboles de zonas templadas pero usuales en árboles tropicales o de pantanos. Ejemplos son Ficus rubiginosa, Ficus elastica, y Ceiba, el género del palo borracho. Las raíces tabulares mantienen el árbol erecto ante los vientos o empujones y lo anclan donde los suelos son poco aptos para enraizar, otorgando alrededor del doble de anclaje extra.[28]
- Raíces tabulares
-
 Raíces tabulares, en este caso con aletones bien conspicuos.
Raíces tabulares, en este caso con aletones bien conspicuos. -
 Raíces tabulares, en este caso los aletones son poco elevados.
Raíces tabulares, en este caso los aletones son poco elevados. -
 Raíces tabulares. Se diferencian de las raíces fúlcreas porque presentan aletones, aunque no se alejen mucho del tronco.
Raíces tabulares. Se diferencian de las raíces fúlcreas porque presentan aletones, aunque no se alejen mucho del tronco. -
 Un ejemplo más modesto de raíces tabulares.
Un ejemplo más modesto de raíces tabulares.





Los pantanos, las zonas bajas e inundables como las palustres, y las marismas, cubren a las raíces de agua impidiéndoles una oxigenación adecuada. Una adaptación en las raíces es llamada neumatóforos, pneumatóforos, neumorrizas o raíces respiratorias, que son raíces que se originan bajo tierra pero crecen hacia arriba, hasta alcanzar el aire (poseen "geotropismo negativo"), y poseen lenticelas en su superficie aérea. Ejemplos: Ludwigia peploides, o el manglar negro, Avicennia germinans.
- Duración
La duración refiere a la longevidad de una planta o de una parte de ella. La duración de la planta se clasifica como anual, bienal o perenne. Una planta anual es una planta que vive por un año o menos, típicamente vive durante la estación de crecimiento, y siempre son hierbas, ya que el lapso de tiempo en el que viven no alcanza para la formación de partes leñosas. Las hierbas anuales pueden usualmente ser detectadas porque carecen de un órgano reservante subterráneo y no muestran evidencias de crecimiento desde una estación previa (por ejemplo, no hay tallos engrosados y leñosos, ni yemas durmientes, ni frutos antiguos) y muestran signos de que van a reproducirse antes de que llegue la estación desfavorable. Las plantas bienales son las que viven por 2 años (o dos estaciones de crecimiento) y usualmente florecen en el segundo año. Las plantas bienales típicamente forman una roseta basal de hojas durante el primer año y florecen al segundo año, y son difíciles de detectar sin observar la misma planta por dos estaciones de crecimiento. Una planta perenne es una planta que vive por más de 2 años. Las perennes pueden ser hierbas con órganos reservantes subterráneos (geófitas), lianas (guiadoras leñosas), hierbas o guiadoras siempreverdes, arbustos, o árboles. Una planta perenne se espera que se desarrolle durante un número de temporadas hasta que alcance la madurez reproductiva. El término monocárpico hace referencia a una planta que florece y da frutos solo una vez, y luego muere, la planta misma puede ser anual o perenne, pero el término es usualmente usado solo para las perennes (porque todas las anuales son monocárpicas por definición). Las plantas que viven más de una estación de crecimiento en ambientes con época desfavorable predecible y que presentan la estrategia ecológica de perder sus partes verdes y rebrotar al comienzo de la nueva estación de crecimiento, poseen siempre para ello órganos reservantes, partes especializadas en almacenar nutrientes utilizados para rebrotar. Pueden provenir del vástago o la raíz, y evolucionaron varias veces independientemente. Las plantas perennes de clima estacional o de época desfavorable predecible que la sobreviven como siempreverdes, presentarán en su hábito caracteres en los que queda expuesta su adaptación a esa estación, como las hojas adelgazadas y coriáceas (de aspecto de cuero) de las coníferas. Son hierbas anuales efímeras las que poseen un ciclo de vida muy corto (en comparación a la duración de una estación favorable) durante el cual se reproducen y mueren, y sobreviven casi todo el año como semilla, son plantas que germinan oportunísticamente luego de una lluvia en los ambientes desérticos.[25]
- Adaptaciones al ambiente (zonas adaptativas)
Síndromes que implican rangos de temperatura y humedad u otras condiciones ambientales incluyen:
- Xeromórfica ("morfología de ambientes secos"), xerófita ("planta de ambientes secos") o xerófila ("amante de ambientes secos") (en ambientes xéricos) si se desarrolla en ambientes secos. Muchas especies epífitas (que crecen sobre otras plantas sin parasitarlas) desarrollan caracteres xeromórficos debido a la velocidad a la que se escurre y se evapora el agua en relación con un sustrato terrestre, así como desarrollan caracteres xeromórficos las plantas de ambientes muy fríos, donde el agua se congela dejando de estar disponible en forma líquida, aprovechable para la planta. Dos adaptaciones xeromórficas son por ejemplo, las raíces muy profundas que toman agua de las napas subterráneas, y las hojas adelgazadas y de cutícula gruesa que previenen la transpiración.[25] Típicamente también lo son los órganos suculentos.
- Higromórfica, higrófita o higrófila (en ambientes húmedos). No confundir con hidrófita (acuática).[29] Es la que se desarrolla en ambientes muy húmedos.[25]
- Una planta mesófita o mesófila (en ambientes mésicos) se desarrolla en ambientes medios de temperatura y humedad.[25]
- Las especies halófitas solamente viven en sustratos que presentan un rango pequeño de altas concentraciones de sal.[25]
- Una de las adaptaciones a un ambiente con variaciones estacionales de temperatura y humedad, tanto de zonas templadas como tropicales, es un tropófito o tropófilo si son las especies de hojas deciduas en período frío o seco, o caducifolias. Otras son las mencionadas geófitas con yemas de renuevo bajo tierra, y las terófitas o anuales, entre las que están las efímeras.
- De cada factor ambiental, cada especie tiene un mínimo y un máximo entre los cuales puede desarrollarse (estos límites también dependen de los valores en otros factores). La distancia entre los dos límites es el rango de tolerancia de la especie. Una especie que sobrevive y se reproduce en un rango amplio de los factores ambientales (con un nicho ecológico amplio) es una especie eurioica.[25] (de lo contrario son estenoicas, adaptadas a vivir en un rango estrecho de una condición extrema.[25])
- Hábitat
Puede indicarse el hábitat:
- terrestre (creciendo sobre la tierra, y si es así si el suelo es arenoso, limoso, arcilloso, con grava, o rocoso),
- epífita (creciendo sobre otra planta, solo usándola como sustrato, sin parasitarla) vista arriba en hábitos,
- saxícola (que crece sobre las rocas o en grietas entre las rocas),
- acuática o hidrófita (creciendo en el agua),
- sumergida (bajo el agua),
- flotante (en la superficie del agua),
- emergente (anclada en el sustrato del fondo con tallos aéreos sobre el agua),
- Parásitas , parásitas epífitas y parásitas epirrizas fueron vistas arriba en hábitos.
- Hábitats
-
 Los "pastos marinos" (que evolucionaron dentro de una serie de familias cercanamente emparentadas de Alismatales: Zosteraceae, Cymodoceaceae, Ruppiaceae y Posidoniaceae) son un ejemplo de plantas acuáticas sumergidas.
Los "pastos marinos" (que evolucionaron dentro de una serie de familias cercanamente emparentadas de Alismatales: Zosteraceae, Cymodoceaceae, Ruppiaceae y Posidoniaceae) son un ejemplo de plantas acuáticas sumergidas. -
 Las "lentejas de agua" (taxón Lemnoideae) son plantas acuáticas flotantes.
Las "lentejas de agua" (taxón Lemnoideae) son plantas acuáticas flotantes. -
 Las marsileáceas (Marsileaceae), entre las que se encuentran los falsos tréboles de 4 hojas, son plantas acuáticas emergentes (palustres).
Las marsileáceas (Marsileaceae), entre las que se encuentran los falsos tréboles de 4 hojas, son plantas acuáticas emergentes (palustres).


.jpg)
-
 Las potamoguetonáceas (Potamogetonaceae) son plantas acuáticas arraigadas con hojas flotantes.
Las potamoguetonáceas (Potamogetonaceae) son plantas acuáticas arraigadas con hojas flotantes. -
 Una Nepenthes saxícola.
Una Nepenthes saxícola.


La descripción del hábitat se completa con la pendiente, el aspecto, la elevación, el régimen de humedad, la vegetación que la rodea, la comunidad, y el ecosistema.
- Distribución
Dos términos contrapuestos relacionados con la distribución son cosmopolita y endémico. Una planta puede ser:
- cosmopolita, si está distribuida en todo el planeta (por lo que debe poseer un nicho ecológico muy amplio, al menos en lo que respecta a la temperatura),
- o endémica si vive en una región geográfica definida. Una especie puede ser endémica aunque su nicho ecológico tolere otra distribución, esto es porque los endemismos son el resultado de, además de las limitaciones del nicho, factores históricos relacionados con el lugar donde la especie se originó, su capacidad de dispersarse, y cambios históricos en el ambiente. Cuando se dice que una planta es endémica se debe especificar endémica a qué zona geográfica es. Nótese que endémica no es sinónimo de estenoica.
- Véase también: Esta sección se completa con los artículos Órganos reservantes en las plantas, Planta de guía, Espinas y aguijones (Botánica).
Raíces
editar- Artículo introductorio: Raíz
A las raíces les faltan los nudos y los entrenudos que caracterizan a los tallos, y también las yemas y las cicatrices foliares que pueden encontrarse en ellos, además de diferenciarse de ellos en importantes caracteres anatómicos que aquí no serán mencionados. Las raíces son bastante uniformes en apariencia, y una planta usualmente no puede ser identificada sin sus partes aéreas.[5] Las raíces son útiles sin embargo, en la determinación de si una planta es anual o perenne, y las variaciones en el sistema radical a veces son taxonómicamente significativas.[5]
- Arquitectura
La primera raíz es la radícula del embrión. Si la radícula continúa desarrollándose después de la etapa de embrión, se la llama raíz primaria. Pueden desarrollarse raíces adicionales:
- A las raíces que se desarrollan a partir del cilindro de otras raíces se las llama raíces laterales (la que se origina en la primaria se llama secundaria, la que se origina en una secundaria se llama terciaria, etc.) A diferencia del tallo que se ramifica en patrones útiles taxonómicamente, usualmente las raíces se ramifican irregularmente.[5]
- A las que se originan en un órgano que no es una raíz se las llama adventicias. Son raíces adventicias las que se originan en los nudos de los tallos de plantas rastreras, estoloníferas, y rizomatosas. Algunos autores también llaman raíces adventicias las que se originan en otra raíz cuando ésta sufrió crecimiento secundario y los primordios radicales poseen un origen diferente que en la raíz de crecimiento primario y deben atravesar la corteza.[4]
- En particular las licofitas se ramifican por división del meristema apical de la raíz[5] (condición solo compartida con las raíces coraloides de las cicadáceas).
-
 La primera raíz es la radícula del embrión. Si se sigue desarrollando luego de la germinación, se la denomina raíz primaria.
La primera raíz es la radícula del embrión. Si se sigue desarrollando luego de la germinación, se la denomina raíz primaria. -
 Raíz adventicia originada en un nudo del tallo de una planta estolonífera.
Raíz adventicia originada en un nudo del tallo de una planta estolonífera.


Se llama sistema radical al patrón formado por todas las raíces de la planta.


- Si la raíz primaria se vuelve dominante, creciendo en grosor y profundidad, se la llama raíz principal[1] (taproot, o axonomorfa o pivotante, en un sistema radical alorrizo),[5] (en árboles puede encontrarse como en forma de estaca, ej. Quercus y algunas coníferas[26])
- Si la raíz primaria se pudre pronto y las raíces subsecuentes son adventicias,[1] o si no se pudre pero por su tamaño y forma no se puede diferenciar de las raíces que la acompañan,[5] teniendo todas las raíces un aspecto homogéneo, ramificadas o no, la planta tiene un sistema radical fibroso u homorrizo
- (En árboles pueden encontrarse raíces en forma de plato, estos casos presentan raíces horizontales superficiales, a partir de las cuales se originan raíces más o menos verticales, ej. Abies, Fraxinus, Populus[26]).

?Averiguar. En algunos árboles y lianas perennes, las raíces también pueden funcionar en una forma de dispersión vegetativa o clonal, cuando son raíces gemíferas, es decir que portan yemas adventicias (embriones radicales?) de las que emergen vástagos nuevos a veces a unos cuantos metros del tronco en el que se originó la raíz. Esta forma de dispersión a corta escala puede generar parches de hábitat donde el árbol con esta forma de propagación sea una especie bien representada.[30] Las especies de Populus (álamos) y Salix (sauces) que crecen a lo largo de la orilla de los ríos, están adaptadas a rebrotar desde sus raíces gemíferas luego de que su tronco principal fuera arrancado por una inundación, o talado por un castor.[30] Otros ejemplos de raíces gemíferas se encuentran en las hierbas Linaria vulgaris y Rumex acetosella.[26]
- Otros tipos de raíces
- Tres tipos de raíces especializados en el sostén y visibles desde la superficie fueron vistas en #Hábito. Las "raíces fúlcreas" o "zancudas" o "zancos", las "raíces tabulares" o "tablares" con "tablas" o "aletones", y las "raíces estranguladoras" de las plantas estranguladoras.
- La función de los órganos reservantes fue indicada en #Hábito. La terminología en las raíces ("raíz tuberosa", "raíz napiforme", ?"caulinotuberosa", ?"xilopodio") en su propio artículo, Órganos reservantes en las plantas.
- Las "raíces aéreas" de las plantas epífitas en #Hábito.
- Los "neumatóforos" en #Hábito.
- "Raíces contráctiles" en el artículo Órganos reservantes en las plantas.
- Las "raíces haustoriales" de las plantas parásitas en #Hábito.
- Las raíces además pueden poseer nódulos, como en la soja, contenedores de una bacteria simbionte fijadora de nitrógeno.
- Se llaman raíces proteoides, raíces proteiformes o raíces en racimo a los densos conglomerados de raíces, descriptos por primera vez en Proteaceae, encontrados en especies que crecen en suelos pobres en nutrientes. Funcionan probablemente modificando químicamente el suelo para mejorar la solubilidad y absorción de los nutrientes. Se las encuentra en plantas adaptadas a sitios pobres en nutrientes (en especial fósforo) y colonizadoras de ambientes xéricos.[31][32]
- En todas las Cycadaceae estudiadas, y sólo en ellas, se encuentran raíces coraloides. Son raíces laterales que se originan a partir de otras raíces expuestas sobre la superficie del suelo, y se extienden sobre él ramificándose dicotómicamente, a veces ingresando hasta 30 cm de profundidad dentro de él. Poseen una estructura celular particular por la que son susceptibles de ser infectadas por el alga simbionte Nostoc, que aparentemente ingresa desde el suelo, y provee a la planta de nitrógeno disponible que fijó desde la atmósfera.[33][34]
- Otras adaptaciones de las raíces
-
 Nódulos en raíz de Lotus pedunculatus.
Nódulos en raíz de Lotus pedunculatus. -

-
 Raíces coraloides infectadas con Nostoc en Cycas revoluta.
Raíces coraloides infectadas con Nostoc en Cycas revoluta.



Tallos y vástagos
editarVer en la introducción.
- Patrón de ramificación del tallo (arquitectura)
Muchos modelos diferentes de patrones de ramificación del tallo (patrón de crecimiento, arquitectura) han sido descritos (ver por ejemplo Hallé et al. 1978[35] citado en Simpson 2005[1] y en Judd et al. 2007[5]).


Una característica principal del patrón de ramificación tiene que ver con la duración del crecimiento del meristema apical de un vástago. Si un vástago en particular tiene potencial para crecer ilimitadamente, de forma que el meristema apical está activo continuamente, el crecimiento se llama indeterminado. Si en lugar de eso el vástago termina su crecimiento después de un período de tiempo, mediante el aborto del meristema apical o su agotamiento en una flor, inflorescencia u otra estructura especializada (una espina caulinar, un zarcillo), entonces el crecimiento es determinado. Estos mismos términos se usan para describir la ramificación de la inflorescencia. Dos términos relacionados,, poco usados: un vástago que se agota en una flor o inflorescencia se llama hapaxántico. Un vástago que porta flores laterales pero continúa su crecimiento vegetativo se llama pleonántico.
Para definir "monopodial" y "simpodial" primero hay que diferenciar la terminología de la morfología del tallo, de la de su patrón de ramificación. En la semilla están diferenciados la plúmula y la radícula, que al germinar serán el tallo primario y la raíz primaria, respectivamente. Esos son el tallo y raíz de primer orden. Si el tallo sigue creciendo y adquiere un grosor y tamaño más importantes que los demás tallos originados en él, coincidirá también con el tallo principal o eje principal. En el tallo primario o de primer orden se originan tallos secundarios o de segundo orden, en ellos los tallos terciarios o de tercer orden, etc. Pero un "eje principal" no siempre está formado por un solo orden de tallos. A veces está formado por unidades, segmentos simpodiales, simpodios, de orden sucesivamente más alto.
Si el eje principal se deriva de un único meristema apical, el patrón de ramificación es monopodial. Cuando el patrón es monopodial, las ramas axilares, las que se originan en las yemas axilares, son también las ramas laterales. El eje monopodial puede crecer indefinidamente y por lo tanto ser indeterminado, dando ramas laterales por desarrollo de las yemas en las axilas de las hojas. De otra manera se llama simpodial. El eje principal está formado por tallos de orden sucesivamente más alto, originados en yemas axilares próximas al meristema apical del tallo de orden anterior, que no sigue formando parte del eje principal. Por ejemplo muchos rizomas son de crecimiento simpodial. El crecimiento simpodial se ve claramente en el seguimiento del desarrollo del tallo de las aráceas terrestres de tallo aéreo y grande, como Philodendron. Finalmente, un tipo raro de ramificación es el dicotómico, en el cual un único meristema apical se divide en dos ramas iguales.
Las plantas monocaules ("un tallo") no se ramifican excepto en la inflorescencia, como en Agave. En las plantas pluricaules ("muchos tallos") el vástago se ramifica.[36]
El tallo o rama puede estar erecto (erguido y además recto, perpendicular al suelo), entonces se llama ortótropo. Cuando se posiciona horizontalmente al suelo se llama plagiótropo.[5][37]
En las imágenes 3 ejemplos de cómo se describe el patrón de crecimiento de una planta.[5]
- Ejemplos de arquitecturas
-
 El eje principal es ortótropo y monopodial, las ramas laterales son plagiótropas y monopodiales. Ejemplos: Duabanga (Lythraceae), Araucaria (Araucariaceae).
El eje principal es ortótropo y monopodial, las ramas laterales son plagiótropas y monopodiales. Ejemplos: Duabanga (Lythraceae), Araucaria (Araucariaceae). -
 El eje principal es ortótropo y monopodial, las ramas laterales son plagiótropas y simpodiales. Ejemplos: Terminalia y Bucida (Combretaceae).
El eje principal es ortótropo y monopodial, las ramas laterales son plagiótropas y simpodiales. Ejemplos: Terminalia y Bucida (Combretaceae). -
 El eje principal es ortótropo y simpodial, las ramas laterales son ortótropas y simpodiales. Ejemplos: Rhus (Anacardiaceae), Pieris (Ericaceae).
El eje principal es ortótropo y simpodial, las ramas laterales son ortótropas y simpodiales. Ejemplos: Rhus (Anacardiaceae), Pieris (Ericaceae).



La descripción de la arquitectura puede incluir la presencia de tipos particulares de tallos con base en su función:
- Tipos de tallo con base en su función
- Órganos reservantes y suculentos indicados en #Hábito. "Bulbo", "cormo", "cáudice", "rizoma", "tubérculo", "bulbillo", "cormillo", "tallo suculento", "tallo caudiciforme", "paquicaulo", "lignotúber" o "lignotubérculo", "pseudobulbo", en su propio artículo, Órganos reservantes en las plantas.
- Estolón indicado en #Hábito.
- Un cladodio o filóclado en #Hábito.
- Las espinas caulinares en Espinas y aguijones (Botánica).
- Un escapo en plantas acaulescentes en #Hábito.
- Un braquiblasto o fascículo en #Hábito.
- Zarcillo caulinar junto con los demás órganos de fijación en Planta de guía.
- Cactaceae. Areola en #Glosario de Cactaceae al final de este artículo.
- Poaceae. Culmo y macollo en #Glosario de Poaceae al final de este artículo.
Yemas
editar- Artículo introductorio: Yema
_nudo_zarcillo_ramificaci%C3%B3n_tallo_tricomas_yema_floral_masculina_florM01_-detalle_etiquetas.png)
Las yemas son el extremo inmaduro, todavía no funcional, de un vástago, con su respectiva región meristemática apical. Una yema se llama vegetativa si se desarrollará como un vástago vegetativo (o como aclara Judd et al. 2007[5]:56, "si posee sólo hojas embrionarias", por lo que podría dar flores también posteriormente a la formación de la yema, como en la yema vegetativa en la foto de Cucurbita), reproductiva si uno reproductivo[38], floral ("flores embrionarias"[5]:56) o inflorescente si flor o inflorescencia respectivamente; o pueden llamarla mixta si posee "flores embrionarias y hojas embrionarias"[5]:56.
La yema en el extremo del eje es la yema terminal o yema apical. En ciertos casos el crecimiento de las hojas y ramas es tan continuo espacialmente que un corte que divida a la yema, con hojas inmaduras (no funcionales), del resto del tallo con hojas maduras, se vuelve una decisión un poco arbitraria. Las que se encuentran en la axila de las hojas son las yemas axilares (o yemas laterales, pero este término puede ser confuso en plantas de crecimiento simpodial). Raramente se pueden encontrar en otros órganos de la planta, como el tronco después del crecimiento secundario (y entonces aparecen por entre la corteza), entonces reciben el nombre de yemas adventicias. Una yema pseudoterminal, en relación al eje en el que se origina, se da si el meristema apical original del vástago aborta o se agota por ejemplo madurando en una flor, y una yema axilar cerca del ápice del tallo continúa el crecimiento vegetativo, tomando la función de una yema terminal. Este es el caso de los ejes simpodiales.
Estas ubicaciones pueden enmascararse debido al fenómeno de concaulescencia, en que el tallo se elonga entre la yema y la hoja dejándola un poco separada de ésta, o el fenómeno de recaulescencia, en que se elonga el pecíolo de la hoja llevándose con él a la yema, quedando ésta sobre la hoja, como en Tilia (el tilo).[39]
- Otras ubicaciones o enmascarados de las yemas
-
 Concaulescencia en la yema floral.
Concaulescencia en la yema floral. -
 Recaulescencia en la yema floral.
Recaulescencia en la yema floral. -
 El fenómeno de recaulescencia hace parecer que las inflorescencias de Tilia se originan en la hoja.
El fenómeno de recaulescencia hace parecer que las inflorescencias de Tilia se originan en la hoja.
.PNG)
.svg)


En algunas especies pueden presentarse yemas axilares múltiples,[39], cuando más de una yema se forma por [axila de una hoja],[1] como en Gossypium (algodón) y Bambusa.[39]
- Las seriales[39] o superpuestas[1] forman una fila a lo largo del eje del tallo, como en Passiflora caerulea[39] (el mburucuyá o Pasión de Cristo), Lonicera japonica[39] (la madreselva), Bougainvillea spectabilis[39] (la Santa Rita), y Turnera orientalis[39]. (Judd et al. 2007[5]:56 llama "superpuestas" solo a las yemas accesorias por arriba o por debajo de una yema axilar principal).
- Las colaterales[39][1] o adyacentes[39] forman una fila horizontal en la axila de la hoja. Esto se observa bien en Allium sativum[39] (el ajo) donde cada diente es una yema axilar, y en el banano[39], donde cada conjunto de yemas colaterales originará una "mano" de bananas.
- Tanto seriales como colaterales pueden estar diferenciadas en una yema principal y yemas accesorias.[5]

Se llama caulifloria a la aparición tardía de flores o inflorescencias en una rama con crecimiento secundario. ([4]:240-241) ?Éstas pueden ser yemas adventicias generadas en forma similar a las raíces endógenas ([4]:240-241).
Si las yemas presentan un período de dormición durante el cual están protegidas por escamas (hojas con aspecto escamoso, pequeñas y no verdes) dispuestas apretadamente, cuando eventualmente la rama crece, estas escamas caen quedando las cicatrices en la base de la rama nueva. Estas ramas se llaman prolépticas y son comunes en especies de clima templado,[5] en las plantas leñosas deciduas, donde las hojas caen al final de la estación de crecimiento.[39] O las yemas pueden desarrollarse y elongarse al mismo tiempo que la rama en la que se originan, en este caso normalmente la rama nueva no tiene escamas basales y tiene un primer entrenudo elongado, estas ramas se llaman silépticas y son comunes en plantas de clima tropical.[5] Algunas plantas, como muchas Lauraceae, tienen tanto ramas prolépticas como ramas silépticas.[5] Son yemas escamosas cuando el ápice está protegido por escamas y yemas desnudas si no poseen estructuras exclusivas de protección, aunque pueden estar algo protegidas por los primordios de las hojas más antiguos y grandes de la yema.[39] Otra estructura de protección presentan las yemas infrapeciolares, que son completamente rodeadas por la base del pecíolo, que también dejará su cicatriz al caer.
- Ramas prolépticas y silépticas
-
 Yemas formadoras de ramas prolépticas.
Yemas formadoras de ramas prolépticas. -
 Las yemas desnudas de esta planta anual forman ramas silépticas.
Las yemas desnudas de esta planta anual forman ramas silépticas.

_new_female_flower_bud_and_tendrils_-_Rama_de_zapallo_con_etiquetas-01.svg)
Corteza (morfología)
editar- Artículo introductorio: Corteza
En el campo de la morfología, cuando se habla de corteza (bark en inglés, no "cortex" en inglés que también puede traducirse como corteza) se hace referencia al aspecto visual de los tejidos protectores más externos de tallos o raíces de las plantas que posean alguna suerte de leñosidad (endurecimiento de los tejidos) en el tallo, sea esta leñosidad derivada de un cámbium suberógeno típico, de un crecimiento secundario anómalo que incluya leñosidad, o de la lignificación (depósitos de lignina en la pared celular) de las células más externas de un crecimiento primario, como en las palmeras y otros árboles con "gigantismo primario".[40] En árboles, la morfología de la corteza muchas veces es un buen carácter para identificar el taxón, particularmente en árboles deciduos durante la estación en que las hojas han caído. Los tipos de corteza, en sentido morfológico, incluyen:
- Exfoliante (exfoliating, peeling[4]), una corteza que se rompe en láminas de aspecto papiráceo (como hojas),
- Fisurada (fissured), la corteza se rompe en muescas verticales u horizontales,
- En placas (plated), una corteza que se rompe en placas aplanadas, más robustas que las papiráceas,
- En hebras, Toscamente fibrosa (shreddy, "bark coarsely fibrous"),
- Lisa (smooth), una corteza no fibrosa (non-fibrous) sin fisuras, fibras (fibers), placas, ni láminas exfoliantes. Sería una corteza sin rugosidades, ver diferencia con inerme.
- Corteza (morfología)
-
 Corteza exfoliante en el "arce de papel" Acer griseum.
Corteza exfoliante en el "arce de papel" Acer griseum. -
 Corteza fisurada.
Corteza fisurada. -
 Corteza en placas en Arbutus unedo.
Corteza en placas en Arbutus unedo. -
![En la corteza lisa se ven más claramente las lenticelas,[4] como en Prunus maackii[4].](//upload.wikimedia.org/wikipedia/commons/thumb/8/8d/Prunus_maackii_01_trunk_by_Line1.jpg/120px-Prunus_maackii_01_trunk_by_Line1.jpg)


.jpg)
![En la corteza lisa se ven más claramente las lenticelas,[4] como en Prunus maackii[4].](/wiki/Archivo:Prunus_maackii_01_trunk_by_Line1.jpg)
La corteza puede estar armada de espinas o aguijones (si no, inerme, comparar con "lisa"). Las espinas y aguijones están descriptos en Espinas y aguijones (Botánica).
Hojas
editar- Artículo introductorio: Hoja
- Artículo principal: Morfología foliar
Se definió hoja, vaina, pecíolo, lámina, estípulas en la introducción. Se comentó que normalmente son bifaciales, aplanadas adaxial-abaxialmente, antero-posteriormente y dorso-ventralmente.
Una hoja unifacial (Kubitzki ed., Simpson 2005, Judd et al. 2007[5]:56) o isobifacial (APWeb) es doble faz pero las dos caras son indiferenciables, usualmente refiere a las hojas isobilaterales (APWeb), es decir, aplanadas con dos caras a los laterales de la línea media (la línea que cruza el tallo donde se origina la hoja), excepto en la base, donde son usualmente envainadoras. Una hoja "céntrica" (centric) es de sección cilíndrica ("terete"), por ejemplo en Fenestraria de Aizoaceae. Algunas monocotiledóneas pertenecientes a muchas familias tienen hojas unifaciales, notablemente las Iridaceae[1].
-
 Hojas unifaciales, isobilaterales, en una iridácea, Iris pumila.
Hojas unifaciales, isobilaterales, en una iridácea, Iris pumila. -
 Hojas céntricas o terete (las verdes, gruesas y suculentas) en Fenestraria, una aizoácea.
Hojas céntricas o terete (las verdes, gruesas y suculentas) en Fenestraria, una aizoácea.


- Partes de la hoja

Algunos autores llaman pseudopecíolo al pecíolo entre vaina y lámina de muchas monocotiledóneas, como Zingiberales (p.ej. Musaceae, la familia de las bananas) y las poáceas bambusóideas.
Si las estípulas están presentes, las hojas son estipuladas. La terminología (opositifolias, axilares o intrapeciolares, ócrea, interpeciolares) en su artículo, Estípula. Las estípulas pueden tener morfologías variadas (foliosas[41], espinas, glándulas, etc) y parece que no siempre son homólogas.
- Las estipelas son un par de estructuras foliosas a cada lado de la base de un foliolo en una hoja compuesta, que también pueden ser modificadas en espinas o glándulas, como en algunas Fabaceae. Si las estipelas están presentes, las hojas son estipeladas.

- Hástula, un apéndice o proyección en la unión del pecíolo y la lámina, como en algunas palmeras.

- Lígula en Poaceae (no en todas), las plantas graminiformes no poáceas no tienen lígula. Una expansión o proyección de la parte superior de la vaina, en su unión con la lámina,.
- Pulvino, la base algo hinchada del pecíolo, o pulvínulo, base algo hinchada del peciolulo; como en algunas Fabaceae. A veces el pulvino o el pulvínulo se encuentran en el ápice del pecíolo (como en Marantaceae) o del peciolulo. A veces el pulvino se encuentra en el medio del pecíolo, como en unas pocas Araceae.[5] El pulvino y el pulvínulo están relacionados con funciones de movimiento de las hojas.[5] El pulvínulo puede, en algunos taxones, funcionar en el movimiento de cierre de los folíolos de una hoja compuesta como respuesta al contacto o al calor (como en Mimosa pudica), también hay una respuesta similar debida al fotoperíodo. Estas respuestas fisiológicas pueden proteger a la hoja del daño mecánico o ayudar a inhibir la pérdida de agua debido a la transpiración. Esta respuesta fisiológica se llama nastia.
- Pulvinos y pulvínulos
-
 Folíolos de Mimosa pudica cerrándose con ayuda de sus pulvínulos, en respuesta al contacto.
Folíolos de Mimosa pudica cerrándose con ayuda de sus pulvínulos, en respuesta al contacto. -
 Hojas de Maranta elevándose con ayuda de sus pulvinos, en respuesta al fotoperíodo.
Hojas de Maranta elevándose con ayuda de sus pulvinos, en respuesta al fotoperíodo. -
 Aspecto de los pulvinos en el pecíolo de Tachigali densiflora.
Aspecto de los pulvinos en el pecíolo de Tachigali densiflora.


_L._F._Gomes_da_Silva_%26_H._C._Lima.jpg)

- Hidátodos, estructuras para eliminar agua activamente por un fenómeno llamado gutación, presentes en plantas que viven en ambientes higrofíticos.[29]
- Domatia y agallas, glándulas. Las estructuras que pueden asociar las hojas con artrópodos pueden ser los "domatia" y las glándulas. Los domatia (literalmente "casitas", singular domatium) o foveolas pueden contener organismos en su interior, usualmente ácaros u hormigas. Los domatia se encuentran en las hojas de muchas angiospermas (Pemberton y Turner 1989,[42] Brouwer y Clifford 1990[43]). Los habitantes artrópodos de los domatia ayudan a la planta a ahuyentar a los herbívoros, y en recompensa, la planta les provee no solo un hogar sino también a veces también alimento. Los domatia de las hormigas son usualmente como un bolsillo y típicamente se encuentran en la base de la lámina de las hojas. Los domatia de los ácaros son más pequeños, usualmente se encuentran en la unión entre las venas de la lámina, pueden tener forma de bol, de volcán, o de bolsillo, pueden estar formados por un grupo de pelos axilares o por un margen revoluto.[5] No se deben confundir los domatia con las agallas, abultamientos en los que se observan insectos alojados, las agallas no son producidas espontáneamente por la planta sino como consecuencia de la invasión por un insecto parásito.
- Domatia y agallas
-
 Domatia para hormigas en Tococa, se encuentran en la base de la hoja, son una estructura hinchada.
Domatia para hormigas en Tococa, se encuentran en la base de la hoja, son una estructura hinchada. -
 Domatium para hormigas en Cordia nodosa, una conspicua hinchazón.
Domatium para hormigas en Cordia nodosa, una conspicua hinchazón. -
 4 domatia para ácaros en las axilas de las venas principales de la lámina de la hoja de Endiandra discolor.
4 domatia para ácaros en las axilas de las venas principales de la lámina de la hoja de Endiandra discolor.



-
 Domatia formados por un grupo de pelos axilares, para ácaros, en el reverso de la lámina de la hoja en Liquidambar styraciflua.
Domatia formados por un grupo de pelos axilares, para ácaros, en el reverso de la lámina de la hoja en Liquidambar styraciflua. -
 Compárese con estas agallas en Eucaliptus, formadas como consecuencia de la invasión de un insecto que lo está parasitando.
Compárese con estas agallas en Eucaliptus, formadas como consecuencia de la invasión de un insecto que lo está parasitando.


- Varias otras estructuras glandulares también pueden aparecer en las hojas. Estas estructuras usualmente secretan néctar y atraen a las hormigas, se llaman nectarios extraflorales, las hormigas a cambio protegen a la planta contra la herbivoría (Bentley 1977[44]).[5] Hay nectarios extraflorales que están relacionados con la polinización en lugar de con la protección por hormigas, como los que se encuentran cercanos a la flor pero no en ella.
- Tipos de hojas según su estructura
El término escama u hoja escamosa es usado para las hojas pequeñas y no verdes (si protegen al meristema apical se llaman pérulas). Una escama rudimentaria encontrada en usualmente plántulas hipógeas se llama catáfilo.
Algunas hojas como brácteas se encuentran en grupos taxonómicos específicos y tienen nombres específicos.
Un filodio es una hoja que consiste en un pecíolo aplanado, como una lámina. Se encuentran en un grupo de especies de Acacia mayormente australianas, y son derivados de hojas ancestralmente compuestas que perdieron el raquis y los folíolos.
Ver zarcillo foliar junto con las demás estructuras de fijación de las guiadoras en Planta de guía.
Tipos de hojas según su patrón de división
editar- Ampliación en Terminología descriptiva de las hojas (Botánica)
El patrón de división de las hojas en componentes discretos o segmentos se llama tipo de hoja. Una hoja simple es la que lleva una única lámina continua. Una hoja compuesta es la que está dividida en dos o más folíolos discretos. El folíolo se diferencia de una hoja simple en que no posee yema en su axila. El tipo de hoja no debe ser confundido con la división de la hoja: la lámina puede estar altamente dividida, pero si no forma folíolos discretos, es técnicamente una hoja simple. En las hojas divididas de los helechos, la primera división es la pina, las siguientes se llaman pínulas. Si las hojas están divididas en dos o más órdenes, se pueden usar también los términos "pina primaria", "pina secundaria", etcétera, con la última división siempre llamándose pínula.
- Hoja simple y compuesta
-
 Hoja simple, se observan la yema, el pecíolo y la lámina.
Hoja simple, se observan la yema, el pecíolo y la lámina. -
 Hoja compuesta, en este caso pinada. Se observa la yema, el pecíolo (en verde oscuro) y el raquis (en verde claro) en el que se insertan los folíolos con su peciolulo (en verde claro).
Hoja compuesta, en este caso pinada. Se observa la yema, el pecíolo (en verde oscuro) y el raquis (en verde claro) en el que se insertan los folíolos con su peciolulo (en verde claro). -
 Hoja compuesta, en este caso palmada. Se observa la yema, el pecíolo (en verde oscuro) y los folíolos con su peciolulo (en verde claro).
Hoja compuesta, en este caso palmada. Se observa la yema, el pecíolo (en verde oscuro) y los folíolos con su peciolulo (en verde claro).



Las hojas simples fueron la condición ancestral en las plantas vasculares, como en los licofilos de las licofitas. También son la norma entre las equisetofitas, psilotofitas, Ginkgo y coníferas y gnétidas. Las hojas compuestas son la norma en los helechos y las cícadas. Hace falta una galería para mostrar cada una. Las angiospermas tienen una gran diversidad de hojas, desde simples a altamente compuestas.
Han evolucionado muchos tipos de hojas compuestas, quizás como una forma de aumentar el área total de la lámina sin sacrificar integridad estructural. Por ejemplo, el tejido de la lámina de una hoja compuesta en general puede tener mejor soporte (por ejemplo en condiciones de viento) que una hoja simple del mismo tamaño. Las hojas compuestas tienden a ser más comunes en ambientes mésicos a húmedos y las hojas simples en ambientes secos, pero hay muchas excepciones y no hay patrones claros.
El eje central de una hoja bipinada ("dos veces pinada") todavía se llama raquis, los ejes laterales que llevan los folíolos se llaman raquillas. Un tipo de hoja costapalmada es la que es esencialmente palmadamente compuesta pero tiene una extensión del pecíolo elongada como si fuera un raquis (llamado costa) como en algunas palmeras.
- Raquilla y Costa
-
 Hoja bipinadamente compuesta. En verde oscuro el pecíolo, en verde claro el raquis, las 6 raquillas y los peciolulos.
Hoja bipinadamente compuesta. En verde oscuro el pecíolo, en verde claro el raquis, las 6 raquillas y los peciolulos. -
 Hoja costapalmada. La parte engrosada en la que se insertan los folíolos es la costa.
Hoja costapalmada. La parte engrosada en la que se insertan los folíolos es la costa.


Se define "hoja pinadamente compuesta", "hoja pinada", "raquis", "imparipinada", "paripinada", "bipinadamente compuesta", "bipinada", "raquillas", "tripinadamente compuesta", "tripinada", "palmadamente compuesta", "palmada", "costapalmada", "costa", "trifoliolada", "ternadamente compuesta", "ternada", "palmada-ternada", "pinada-ternada", "geminada", "bifoliolada", "bigeminada", "geminada-pinada", "unifoliolada", en Terminología descriptiva de las hojas (Botánica).
Inserción de la hoja en el tallo
editarSe definió pecíolo, peciolulo, folíolo, vaina, envainadora, en la #Introducción a los órganos de la planta.
Una hoja no peciolada o un folíolo no peciolulado es sésil. También es usado el término subsésil para la hoja con un pecíolo corto o el folíolo con un peciolulo corto
- Inserción de la hoja en el tallo
-
Hoja peciolada.
-
 Hoja sésil.
Hoja sésil.

Se define "decurrente", "amplexicaula", "perfoliolada", "conada-perfoliolada", en Terminología descriptiva de las hojas (Botánica).
Venación de las hojas
editarSe definió "venas" o "nervios" (y por lo tanto "venación" o "nerviación") en la #Introducción a los órganos de la planta.

La vena que se continúa con el pecíolo es la vena primaria. De la vena primaria pueden ramificarse venas secundarias, de las venas secundarias pueden ramificarse venas terciarias, etc. La vena primaria también puede llamarse vena media o costa ("costa" tiene otra definición en briofitos).
Cuando en el pecíolo o cerca de él se originan más de una vena con el mismo grosor, se llaman venas principales. Algunos autores pueden llamarlas venas primarias también.
Los patrones de venación pueden ser muy complejos, y la terminología enorme. Se diferencia particularmente la nerviación paralela (los haces primarios paralelos) de la reticulada (haces en red), que son los dos tipos de nerviación ancestrales de monocotiledóneas y dicotiledóneas, respectivamente. En monocotiledóneas, la nerviación paralela coincide con una base foliar envainadora que toma las venas de un radio amplio de la circunferencia del tallo. Las venas secundarias en la venación paralela son típicamente transversales, quedando la nerviación "escalariforme", pero puede tener una venación secundaria reticulada también.
Los términos "nerviación abierta", "nerviación cerrada", "uninervia", "nerviación dicotómica", "nerviación paralela", "nerviación reticulada", "areolas", "vénulas", "palmadamente venadas", "pinadamente venadas"; "ternadamente venadas", "venación peni-paralela" o "peniparalela", "venación palmatiparalela", "percurrente"; en Terminología descriptiva de las hojas (Botánica)#Venación de las hojas.
El sistema de Hickey (1973[45]) y Hickey y Wolfe (1975[46]): "craspedódroma", "camptódroma", "hipódroma", "craspedódroma simple", "semicraspedódroma", "craspedódroma mixta", "broquidódroma", "eucamptódroma", "cladódroma", "reticulódroma", "paralelódroma", "actinódroma", "palinactinódroma", "marginal", "reticulada", "flabelada", "campilódroma", "acródroma", "basal", "perfecta", "suprabasal", "imperfecta", en Terminología descriptiva de las hojas (Botánica)#Venación de las hojas.
Forma y márgenes de las hojas
editarLas hojas pueden tener diferentes forma, márgenes, color, aspecto de su superficie, etc. Estos caracteres que también pueden aplicarse a otras partes de la planta se encuentran más adelante, en la sección de "#Terminología general". Márgenes y dientes de la hoja en Terminología descriptiva de las hojas (Botánica)#Márgenes.
Flores
editar- Artículo introductorio: Flor
- Artículo principal: Morfología floral
Como definido en castellano y se enseña en las Universidades hispanas, una flor es un braquiblasto especializado en un síndrome de reproducción, que porta las hojas fértiles (estambres, carpelos). Como definido en castellano, todas las espermatofitas tienen flores. Los "conos" de las gimnospermas son flores o inflorescencias, como también la flor de angiosperma, la que posee el óvulo "envasado" en el carpelo, que después de desarrollarse es la semilla "envasada" en el fruto, órgano que le da nombre al grupo.
- Piezas y partes de la flor


Se definió "flor", "pedicelo", "receptáculo" o "tálamo", "antofilos" o "piezas florales", "sépalos", "pétalos", "perianto", "estambres", "carpelos", "profilos", "antera", "grano de polen", "líneas de dehiscencia", "estilo", "estigma", "óvulo", "ovario", "pistilo", "semilla" en la #Introducción a los órganos de la planta.
La hoja en cuya axila se originó la flor, ubicada en el tallo en el que está inserta, puede ser un nomófilo,[47] pero si difiere en su morfología se la denomina bráctea. La bráctea puede formar parte del "síndrome floral", como en poáceas y varias otras familias del orden Poales en que es una de las brácteas que protege a la flor del paso de los animales caminadores.
Los sépalos en conjunto forman el cáliz, los pétalos en conjunto forman la corola, ambos en conjunto son el perianto o perigonio (los antofilos no reproductivos), que si evolucionaron de forma de tener la misma función petaloide son llamados tépalos. Los estambres en conjunto forman el androceo, y en angiospermas ancestrales presentan dos tecas (cada teca con dos microsporangios) en una sola antera. Es decir que cada antera tiene típicamente 4 microsporangios. Los estambres pueden ser aplanados ("laminares") pero típicamente constan de un filamento que porta la antera en el extremo apical. Los carpelos en conjunto forman el gineceo.
- Partes de la flor
-
 La azucena, Lilium. 1-2: Gineceo 1: Estigma 2: Estilo 3-4: Estambres 3: Anteras 4: Filamentos 5: Tépalos (pétalos y sépalos, indiferenciados).
La azucena, Lilium. 1-2: Gineceo 1: Estigma 2: Estilo 3-4: Estambres 3: Anteras 4: Filamentos 5: Tépalos (pétalos y sépalos, indiferenciados). -
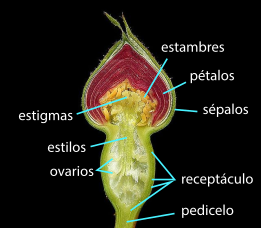 En el pimpollo (yema floral) se ve claramente la función protectora de los sépalos. En la foto un pimpollo de rosa (género Rosa), el receptáculo está expandido hasta formar una cavidad en la cual se desarrollan los pistilos.
En el pimpollo (yema floral) se ve claramente la función protectora de los sépalos. En la foto un pimpollo de rosa (género Rosa), el receptáculo está expandido hasta formar una cavidad en la cual se desarrollan los pistilos. -
 Flor de loto (Nelumbonaceae), se observan los pétalos (los sépalos debajo de ellos no se observan), los estambres, y el resto del receptáculo elevado (verde) dentro del cual se encuentran inmersos los pistilos.
Flor de loto (Nelumbonaceae), se observan los pétalos (los sépalos debajo de ellos no se observan), los estambres, y el resto del receptáculo elevado (verde) dentro del cual se encuentran inmersos los pistilos.



-
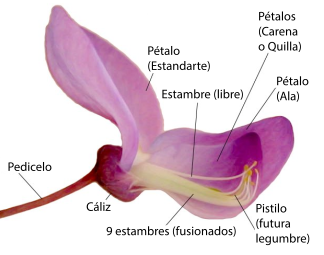 Fusión de algunas piezas: flor de una leguminosa papilionóidea. 5 sépalos conados (fusionados entre sí), 5 pétalos variadamente conados, 10 estambres (9 conados y 1 libre) y un pistilo que se asoma por entre el tubo formado por los estambres conados. (De los pétalos, un ala removida, la quilla está formada por 2 pétalos conados, uno de ellos removido).
Fusión de algunas piezas: flor de una leguminosa papilionóidea. 5 sépalos conados (fusionados entre sí), 5 pétalos variadamente conados, 10 estambres (9 conados y 1 libre) y un pistilo que se asoma por entre el tubo formado por los estambres conados. (De los pétalos, un ala removida, la quilla está formada por 2 pétalos conados, uno de ellos removido). -
 Flor del baobab, Adansonia digitata, una malvácea. Se observan sépalos, pétalos, estambres (filamentos fusionados en la base, anteras marrones) y carpelos (gineceo blancuzco con ovario, estilo y estigma). El estilo del gineceo está doblado 90º, se exerta del tubo formado por los filamentos de los estambres fusionados.
Flor del baobab, Adansonia digitata, una malvácea. Se observan sépalos, pétalos, estambres (filamentos fusionados en la base, anteras marrones) y carpelos (gineceo blancuzco con ovario, estilo y estigma). El estilo del gineceo está doblado 90º, se exerta del tubo formado por los filamentos de los estambres fusionados.
_labels_spanish_c%C3%A1liz_estambres_diadelfos_p%C3%A9talos_quilla_carena_estandarte_alas_pistilo.svg)

Morfologías del receptáculo.
Las piezas pueden estar ubicadas sobre estructuras alargadas que las elevan y que pueden provenir del receptáculo o de las mismas piezas fusionadas. Aquí se dará la terminología del glosario del APWeb.[48]
- ginóforo, el gineceo es elevado por un pie proveniente del receptáculo. El ginóforo no necesariamente es más delgado que el gineceo ubicado sobre él. (compárese con el "gineceo estipitado": el gineceo en su parte inferior presenta una constricción en forma de pie, el estípite o "ginopodio".)
- androginóforo, el gineceo junto con el verticilo de estambres son elevados por un pie proveniente del receptáculo, como en las pasionarias (familia Passifloraceae).
- antóforo, el gineceo, los estambres y los carpelos son elevalos por un pie proveniente del receptáculo, como en Lychnis (foto).
- andróforo, una elevación (normalmente en forma de tubo, aunque no necesariamente) formada por el receptáculo que eleva solo el verticilo de estambres, cuando es en forma de tubo, rodea al gineceo. (compárese con el "tubo estaminal" formado por los "estambres monadelfos", es decir con sus bases fusionadas, conadas, ver en Estambres).
- hipanto, un tubo formado por el receptáculo ("origen axial") o las bases de las piezas fusionadas ("origen apendicular") o una combinación de ambos, que eleva estambres, pétalos y sépalos. Esta definición de hipanto no es utilizada por todos los autores, por eso se puede encontrar como "hipanto libre" (no adnato a la pared de los carpelos de un ovario ínfero).
- disco, las elevaciones del receptáculo intercaladas entre los verticilos de piezas florales en forma de anillo, o que elevan los verticilos internos a ella en un plato. En general los discos son nectaríferos, ver en la sección Nectarios florales.
-
![Ginóforo en Capparaceae, a su alrededor numerosos estambres de filamentos largos.[49]](//upload.wikimedia.org/wikipedia/commons/thumb/1/1e/Capullito_capullito.JPG/244px-Capullito_capullito.JPG) Ginóforo en Capparaceae, a su alrededor numerosos estambres de filamentos largos.[49]
Ginóforo en Capparaceae, a su alrededor numerosos estambres de filamentos largos.[49] -
 Androginóforo en una pasiflorácea (pasionaria, pasión de Cristo o mburucuyá), Passiflora caerulea.
Androginóforo en una pasiflorácea (pasionaria, pasión de Cristo o mburucuyá), Passiflora caerulea. -
![Cleome gynandra presenta andróforo y ginóforo: el andróforo en forma de tubo rodea al ginóforo que porta el gineceo.[50]](//upload.wikimedia.org/wikipedia/commons/thumb/f/ff/Cleome_gynandra_3_%284545384955%29.jpg/260px-Cleome_gynandra_3_%284545384955%29.jpg) Cleome gynandra presenta andróforo y ginóforo: el andróforo en forma de tubo rodea al ginóforo que porta el gineceo.[50]
Cleome gynandra presenta andróforo y ginóforo: el andróforo en forma de tubo rodea al ginóforo que porta el gineceo.[50]
![Ginóforo en Capparaceae, a su alrededor numerosos estambres de filamentos largos.[49]](/wiki/Archivo:Capullito_capullito.JPG)

![Cleome gynandra presenta andróforo y ginóforo: el andróforo en forma de tubo rodea al ginóforo que porta el gineceo.[50]](/wiki/Archivo:Cleome_gynandra_3_(4545384955).jpg)
"Estípite" es una palabra general para referirse a este tipo de estructuras morfológicas, como "pie" y "columna", y si son de poca altura "disco" (un disco puede tener forma de disco o de anillo, con otros verticilos de piezas florales internos a él), sin adjetivar no refieren al tejido del que provienen, aunque por disco se suele referir a los discos provenientes del receptáculo ya mencionados. Estructuras como las anteriores pueden provenir de piezas fusionadas y tienen nombres diferentes que los indicados más arriba, como en el caso del gineceo estipitado o ginopodio ya mencionado, o en los estambres conados (fusionados) por sus filamentos del ejemplo de más arriba, llamados estambres monadelfos (si están fusionados en dos grupos, o en un grupo y un estambre libre como en el ejemplo de las leguminosas papilionóideas, son estambres diadelfos). En algunos taxones el androceo y el gineceo están fusionados en una estructura común llamada ginostemio (Orchidaceae) o ginostegio (Apocynaceae), ambos formadores de "polinarios" (ver en la cita[cita 2] y en sus artículos de las familias respectivas).
-
 ginóforo y gineceo estipitado
ginóforo y gineceo estipitado -
 andróforo y estambres monadelfos
andróforo y estambres monadelfos


La simetría de la flor a veces no es obvia, y puede variar dentro de los verticilos de una flor, si es así, hay que describir la simetría por separado del cáliz, corola, androceo y gineceo. Las flores de simetría radial son actinomórficas ("con forma de estrella", "estrelladas"), en vista distal se dividen en más de dos planos de simetría. Las flores de simetría bilateral en vista distal son las zigomórficas o cigomórficas, las divide un solo plano de simetría, en general coincide con una orientación del eje floral más paralela al suelo (entonces existiendo un lado ventral y un lado dorsal de la flor), y hay muchas formas diferentes en que la zigomorfía puede desarrollarse, habiéndose originado en muchos grupos independientemente. Típicamente evoluciona como una forma de transferir polen más eficientemente a un animal polinizador, usualmente un insecto. Se ven muchos ejemplos más adelante. El giro sobre su eje en 180º de alguna de las partes, de forma que queda "dada vuelta", se llama resupinación, y puede ocurrir también en hojas. En orquídeas se resupina la flor completa por torcedura del pedicelo, pero como cuando esto ocurre no pierde su simetría, siguen siendo flores de simetría bilateral. Si la flor no puede dividirse en ningún plano de simetría es asimétrica. Se definieron: "simetría radial", "flor radiada", "actinomórfica", "regular", "polisimétrica", "birradial", "bilateral", "zigomórfica" o "cigomórfica", "monosimétrica", "cíclica", "asimétrica", "acíclica" en el artículo Terminología descriptiva de las flores (Botánica).
- Simetría floral
-
 Commelina, flor zigomórfica, de simetría bilateral.
Commelina, flor zigomórfica, de simetría bilateral. -
 Una amarilidácea, flor actinomórfica, de simetría radial.
Una amarilidácea, flor actinomórfica, de simetría radial. -
 Flor asimétrica.
Flor asimétrica.



-
![Las cannáceas (achiras) poseen 4 estaminodios petaloideos y un estambre fértil petaloideo, el estambre fértil posee una única teca fértil alargada visible en el centro de la foto (a su lado se ve el estilo, también petaloideo, más pequeño que los estaminodios). (Según Judd et al. 2007[5] el estambre fértil está adnato a un estaminodio petaloideo, por lo que son dos piezas fusionadas -connadas-) Los tépalos son robustos y no petaloideos. El sector del androceo es asimétrico.](//upload.wikimedia.org/wikipedia/commons/thumb/2/20/Canna_-_Howrah_2011-02-20_1536_spanish_labels_t%C3%A9palos_estaminodios_estambre_antera_f%C3%A9rtil_estilo_petaloideo.jpg/337px-Canna_-_Howrah_2011-02-20_1536_spanish_labels_t%C3%A9palos_estaminodios_estambre_antera_f%C3%A9rtil_estilo_petaloideo.jpg) Las cannáceas (achiras) poseen 4 estaminodios petaloideos y un estambre fértil petaloideo, el estambre fértil posee una única teca fértil alargada visible en el centro de la foto (a su lado se ve el estilo, también petaloideo, más pequeño que los estaminodios). (Según Judd et al. 2007[5] el estambre fértil está adnato a un estaminodio petaloideo, por lo que son dos piezas fusionadas -connadas-) Los tépalos son robustos y no petaloideos. El sector del androceo es asimétrico.
Las cannáceas (achiras) poseen 4 estaminodios petaloideos y un estambre fértil petaloideo, el estambre fértil posee una única teca fértil alargada visible en el centro de la foto (a su lado se ve el estilo, también petaloideo, más pequeño que los estaminodios). (Según Judd et al. 2007[5] el estambre fértil está adnato a un estaminodio petaloideo, por lo que son dos piezas fusionadas -connadas-) Los tépalos son robustos y no petaloideos. El sector del androceo es asimétrico.
![Las cannáceas (achiras) poseen 4 estaminodios petaloideos y un estambre fértil petaloideo, el estambre fértil posee una única teca fértil alargada visible en el centro de la foto (a su lado se ve el estilo, también petaloideo, más pequeño que los estaminodios). (Según Judd et al. 2007[5] el estambre fértil está adnato a un estaminodio petaloideo, por lo que son dos piezas fusionadas -connadas-) Los tépalos son robustos y no petaloideos. El sector del androceo es asimétrico.](/wiki/Archivo:Canna_-_Howrah_2011-02-20_1536_spanish_labels_t%C3%A9palos_estaminodios_estambre_antera_f%C3%A9rtil_estilo_petaloideo.jpg)
Si hay más de una pieza por nudo la flor es verticilada. Puede haber solo una pieza por nudo, y cuando es así las piezas se suceden formando una espiral: son flores espiraladas. Estos dos arreglos pueden darse también en sólo un sector de la flor. Típicamente, las plantas con el perianto arreglado en espiral (flores de perianto espiralado, perianto de disposición espiralada) tienen piezas del perianto que son o bien indiferenciadas, o bien que se intergradan una con otra de una forma sepaloide a una forma petaloide. En la mayoría de las angiospermas la disposición es verticilada, no necesariamente en el primordio foliar, pero sí a la madurez.
- Flores espiraladas y verticiladas
-
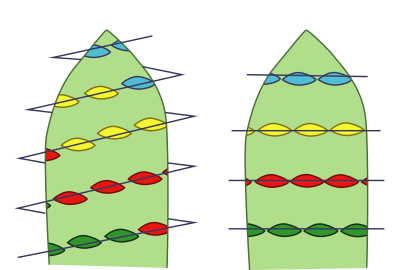 Receptáculo (verde claro) y disposición espiralada y verticilada de las piezas.
Receptáculo (verde claro) y disposición espiralada y verticilada de las piezas. -
 Flor de perianto y androceo verticilados (Alisma plantago-aquatica), 3 sépalos en el primer verticilo, 3 pétalos en el segundo verticilo, piezas de uno y otro verticilo alternadas.
Flor de perianto y androceo verticilados (Alisma plantago-aquatica), 3 sépalos en el primer verticilo, 3 pétalos en el segundo verticilo, piezas de uno y otro verticilo alternadas.


-
 Flor de piezas espiraladas en el camalote del género Nymphaea.
Flor de piezas espiraladas en el camalote del género Nymphaea. -
 Sucesión de morfología de estambres en una ninfácea, en la imagen el paso de pétalos a estambres. El de la izquierda todavía es un pétalo, o puede ser considerado un estaminodio (estambre no funcional), se insinúan los sitios de las tecas de la antera. El resto son estambres laminares (la antera formada por una teca a cada lado del sector apical) y el de la derecha ya puede ser considerado un estambre filamentoso.
Sucesión de morfología de estambres en una ninfácea, en la imagen el paso de pétalos a estambres. El de la izquierda todavía es un pétalo, o puede ser considerado un estaminodio (estambre no funcional), se insinúan los sitios de las tecas de la antera. El resto son estambres laminares (la antera formada por una teca a cada lado del sector apical) y el de la derecha ya puede ser considerado un estambre filamentoso.


Maduración centrífuga se refiere a que la flor madura desde el centro hasta la periferia, mientras que maduración centrípeta se refiere al caso contrario. Los dos términos pueden referirse a partes de la flor, o sólo el gineceo, androceo, o perianto, los términos son muchas veces usados para describir la dirección de maduración de los estambres en un androceo con más de un verticilo. En las inflorescencias, como los girasoles, también se aplican los términos de maduración centrífuga/centrípeta, y se refieren a la dirección de la maduración de las flores individuales dentro de la inflorescencia.
- Maduración centrífuga/centrípeta
-
 Maduración centrípeta de las flores individuales en la inflorescencia de Lantana.
Maduración centrípeta de las flores individuales en la inflorescencia de Lantana. -
 Maduración centrípeta de las flores individuales en la cabeza de un girasol. En los girasoles, cada "pétalo" es una flor y cada "semilla" es una flor.
Maduración centrípeta de las flores individuales en la cabeza de un girasol. En los girasoles, cada "pétalo" es una flor y cada "semilla" es una flor.


Nectarios florales y otras recompensas
editar- Véase también: Nectario
Los nectarios son estructuras especializadas en producir y secretar néctar, si se presentan en la flor son nectarios florales y fuera de la flor son nectarios extraflorales, las estructuras que secretan néctar se llaman necaríferas (es decir son nectarios), y las que lo almacenan, si presentes, colectoras de néctar. El néctar es una solución compleja cuya composición depende de la especie, pero para ser llamada néctar debe tener cierta concentración de azúcares. El néctar funciona como recompensa de la polinización animal, son atraídos por flores que aparentan tener néctar los murciélagos, aves, otros mamíferos, y especialmente insectos (nótese que "atraer" no es sinónimo de "recompensar", a veces los agentes polinizadores pueden ser engañados). A veces los nectarios son tan pequeños o están tan ocultos en los tejidos internos de la planta que sólo son aparentes cuando se encuentra el néctar secretado. Se debe sospechar que la flor es nectarífera cuando se encuentran estructuras que parecen especializadas en colectarlo (p.ej. pétalos con espolones, bolsillos, o piezas ocultando cámaras colectoras) o cuando se encuentra que la flor oculta o desvía la atención del polen al visitante haciéndole presumir que hay otra recompensa (p.ej. flores de estructura bilabiada, de estructura aquillada, ver en #Partes y tipos de perianto (y en general de estructuras periantoideas)).

Según su origen del desarrollo se pueden clasificar en nectarios:
- receptaculares o axiales (formados por el receptáculo)
- es común el disco nectarífero, un crecimiento del receptáculo entre verticilos con forma de plato o si quedan verticilos florales internos, con forma de anillo (algunos autores a este último lo pueden llamar anillo nectarífero). Los glosarios suelen mencionar si el disco es
- intraestaminal (ubicado entre estambres y gineceo),
- estaminal (que porta los estambres) o
- extraestaminal (ubicado entre los estambres y los pétalos).
- es común el disco nectarífero, un crecimiento del receptáculo entre verticilos con forma de plato o si quedan verticilos florales internos, con forma de anillo (algunos autores a este último lo pueden llamar anillo nectarífero). Los glosarios suelen mencionar si el disco es
- phyllome nectaries, formados por una pieza floral.
- Los nectarios septales son tejidos especializados embebidos dentro de los septos del ovario, se observa el néctar secretado a través de un poro en la base del ovario o en el ápice.
- El perigonial está en el perianto, usualmente en la base de los sépalos, de los pétalos, o de los tépalos.
Otras glándulas especializadas pueden secretar compuestos no azucarados, recompensas que no clasifican como néctar. Éstos no se llaman nectarios,Error en la cita: Parámetro inválido en la etiqueta <ref>[51] por ejemplo (más ejemplos en Weberling (1989[51] p. 194):
- las glándulas de la cera que secretan los miembros de KrameriaceaeError en la cita: Parámetro inválido en la etiqueta
<ref> - los elaióforos (estructuras con glándulas secretoras de aceites) en los sépalos de las Malpighiaceae del Nuevo Mundo (Cypella, Diascia, Calceolaria), aceites utilizados por ciertas especies de abejas para alimentar a su cría (Vogel 1974).
La flor también puede recompensar con tejido vegetal en lugar de secreciones:
- los pelos alimenticios (feeding hairs, tricomas que son ellos mismos alimento de insectos) como los que secretan aceites y proteínas en el labelo de Maxillaria,
- los tricomas ricos en azúcar en Sterculia javanica (Cammerloher 1931)
Brácteas de la flor e inflorescencia
editarSe definieron los profilos en la #Introducción a los órganos de la planta.

Pueden encontrarse estructuras foliosas cerca del cáliz que formen parte funcional de la flor. Un grupo de piezas que parecen un verticilo "extra" de sépalos, formado por brácteas o bracteolas inmediatamente debajo del cáliz se llama epicáliz o calículo. Se encuentra por ejemplo en muchos miembros de Malvaceae sensu stricto. Cuando las piezas del epicáliz están soldadas entre sí se denomina gamófilo.
En la misma función que el epicáliz o calículo, pero alejadas del cáliz por quedar debajo de un ovario ínfero, son las brácteas involucrales que justamente, están involucradas formando un involucro (no confundir con el involucro de filarios de las inflorescencias de Asteraceae), como en la familia Fagaceae (la familia de los robles, Quercus, y castaños, Castanea). Algunos pueden presentar brácteas no involucradas en esa posición. El involucro a veces está fusionado formando una cúpula como la que se encuentra en la base y cubriendo parte de las bellotas.[5]:403
- Brácteas involucrales y relativos
-
 Cúpula de brácteas involucrales espinosas (arriba) y castañas (abajo) en Castanea dentata, una fagácea.
Cúpula de brácteas involucrales espinosas (arriba) y castañas (abajo) en Castanea dentata, una fagácea. -

-

-
 El nogal (Juglans regia, Juglandaceae) posee un involucro que rodea la flor femenina y puede desarrollarse formando parte funcional del fruto (ser "acrescente"), hasta formar alas o, como en el caso del nogal, rodeando la nuez en una capa verde carnosa.
El nogal (Juglans regia, Juglandaceae) posee un involucro que rodea la flor femenina y puede desarrollarse formando parte funcional del fruto (ser "acrescente"), hasta formar alas o, como en el caso del nogal, rodeando la nuez en una capa verde carnosa.



_and_female_flowers_spanish_label_involucro.jpg)
Una espata es una bráctea alargada, muchas veces robusta, a veces coloreada, debajo de y usualmente encerrando a la inflorescencia de muchas monocotiledóneas, por ejemplo en Araceae.
- Espatas
-
 Espata en la inflorescencia de Arum palaestinum (Araceae), que protege las flores, muy pequeñas, dispuestas apretadamente alrededor del eje grueso llamado espádice.
Espata en la inflorescencia de Arum palaestinum (Araceae), que protege las flores, muy pequeñas, dispuestas apretadamente alrededor del eje grueso llamado espádice. -
 Espata en Anthurium, otra arácea.
Espata en Anthurium, otra arácea. -
 Espata en la inflorescencia de Strelitzia, las "aves del paraíso" (Strelitziaceae).
Espata en la inflorescencia de Strelitzia, las "aves del paraíso" (Strelitziaceae). -
 Espatas leñosas en la inflorescencia de una palmera (Arecaceae).
Espatas leñosas en la inflorescencia de una palmera (Arecaceae).



_foto_02.JPG)
Asteraceae. Las brácteas en flor e inflorescencia en #Glosario de Asteraceae al final de este artículo.
Poaceae. Las brácteas en flor e inflorescencia en #Glosario de Poaceae al final de este artículo.
Otras definiciones de la flor
editarSexo de la flor y sexo de la planta. Se definió "flor perfecta", "bisexual", "monoclina", "hermafrodita", "imperfecta", "unisexual", "diclina", "carpelada", "estaminada", "incompleta", "neutra", "asexual", "monoica", "dioica", "polígama", "andromonoica", "ginomonoica", "trimonoica", "androdioica", "ginodioica", "trioica", en el artículo Terminología descriptiva de las flores (Botánica).
Inserción de la flor al tallo. Se definió "pedicelada", "sésil", "subsésil", "bracteada", "no bracteada", en el artículo Terminología descriptiva de las flores (Botánica).
Maduración de la flor y la inflorescencia. Se definió "protandria", "protoginia", en el artículo Terminología descriptiva de las flores (Botánica).
Perianto
editarEl perianto o perigonio fue definido en la introducción y en #Flor.
- Partes y tipos de perianto (y en general de estructuras periantoideas)
En esta sección se encontrarán las estructuras formadas por los órganos petaloideas y sepaloideas, provengan del perianto (que es lo más general) o no. Los "tipos de perianto" o más generalmente "construcciones con órganos petaloideas y sepaloideas", típicamente reflejan en un grado importante las adaptaciones relacionadas con la forma en que el polinizador es atraído y la forma en que el polen le es transferido o recibido por parte de él, por lo que es una estructura fundamental para comprender cómo se cumple la función de polinización en cada tipo de flor (véase síndrome floral).
En los tipos de perianto la terminología tiene en cuenta varios aspectos de la forma, fusión, orientación, y merosidad (número de piezas) del perianto. El tipo de perianto y la morfología de las partes del perianto son muchas veces de gran valor taxonómico y puede ser diagnóstico de muchos grupos de angiospermas.
Corona. Lo que pareciera ser un verticilo "extra" de piezas petaloideas ubicado justo por encima del verticilo de pétalos, muchas veces vistoso, es llamado corona o paraperigonio (y son flores "coronadas"), su origen del desarrollo pueden ser las piezas del perianto, el androceo, o el receptáculo.[5] Tienen corona los narcisos (Narcissus) y Asclepias. Pueden llamarse escamas coronales a los sobrecrecimientos en la parte adaxial de los pétalos con función de corona, como en Boraginaceae, y también a lo que algunos autores prefieren llamar "crecimientos ligulados" o lígulas, como ocurre en los pétalos de algunas Gentianaceae y muchas Silenoideae (compárese con la lígula en la hoja de Poaceae) (Weberling 1989[51] p. 80).
- Corona y escamas coronales, lígulas
-
 Narcissus, pétalos blancos y corona crema.
Narcissus, pétalos blancos y corona crema. -
 Escamas coronales en Nerium oleander (adelfa, baladre, laurel de flor).
Escamas coronales en Nerium oleander (adelfa, baladre, laurel de flor). -
 Escamas coronales señaladas en Borago officinalis.
Escamas coronales señaladas en Borago officinalis. -
 Escamas coronales o lígulas en Gentiana verna.
Escamas coronales o lígulas en Gentiana verna.




Unguiculado. El delineado de un pétalo puede ser muy variable, pero vale la pena mencionar que en pétalos libres, la parte basal puede estar adelgazada y bien diferenciada de la parte distal ensanchada y vistosa, llamadas respectivamente uña y limbo. La uña ("claw" en inglés) puede ser muy corta, como en Rosa o muy larga como en el clavel, Dianthus, y normalmente está escondida por el cáliz.[52][51] Un perianto de piezas con uña y limbo (y por lo tanto también dialipétalo, con piezas separadas entre sí) se llama perianto unguiculado (en inglés clawed), presente en numerosas familias, como muchas Brassicaceae (repollo, col) y Caryophyllaceae (la familia de los claveles). El árbol del cacao Theobroma cacao tiene la uña expandida en forma de copa[53], en un capuchón petalar[54] que protege el estambre.
- Uña y capuchón petalar
-
 Perianto unguiculado (pétalos separados entre sí, con uña y limbo).
Perianto unguiculado (pétalos separados entre sí, con uña y limbo). -
 Pétalo de clavel (Dianthus: Caryophyllaceae). Uña blanquecina; limbo coloreado.
Pétalo de clavel (Dianthus: Caryophyllaceae). Uña blanquecina; limbo coloreado. -
 Pétalo de Geraniaceae, uña y limbo.
Pétalo de Geraniaceae, uña y limbo. -
 Flor de Theobroma cacao, las uñas de los pétalos expandidas en capuchones petalares que protegen los estambres.
Flor de Theobroma cacao, las uñas de los pétalos expandidas en capuchones petalares que protegen los estambres.




Cruciado. Las brasicáceas poseen 4 pétalos unguiculados, enfrentados de dos en dos cuya parte distal se refleja abruptamente formando la figura de una cruz en el plano de la vista distal, es el perianto cruciado por el que se las conoce como crucíferas (Cruciferae, el antiguo nombre sinónimo conservado de las brasicáceas).
- Perianto cruciado
-
 Perianto cruciado (4 pétalos unguiculados reflejos formando una cruz).
Perianto cruciado (4 pétalos unguiculados reflejos formando una cruz). -
 Perianto cruciado en una brasicácea o crucífera.
Perianto cruciado en una brasicácea o crucífera.


En un perianto de piezas conadas (fusionadas entre sí) se pueden encontrar: el tubo (tube), la región basal cilíndrica, usualmente de una corola gamopétala (con pétalos fusionados entre sí por sus márgenes) y los lóbulos que son las proyecciones en la parte externa que ya no forman el tubo, la transición puede llamarse garganta (throat). Los lóbulos esperablemente coinciden con la posición en la que está inserta la pieza a la que pertenecen, se encuentran en su mismo radio.
- Tubo, garganta y lóbulos
-
 Una bignoniácea, se observa el tubo ensanchado en una garganta, y el perianto bilabiado (ver más abajo).
Una bignoniácea, se observa el tubo ensanchado en una garganta, y el perianto bilabiado (ver más abajo). -
 El jacarandá (Jacaranda, una bignoniácea), tubo, garganta y lóbulos.
El jacarandá (Jacaranda, una bignoniácea), tubo, garganta y lóbulos. -
 Tabernaemontana divaricata (una apocinácea), tubo, garganta y lóbulos.
Tabernaemontana divaricata (una apocinácea), tubo, garganta y lóbulos. -
 Campanitas de Navidad australianas (familia Blandfordiaceae), tubo, garganta y lóbulos.
Campanitas de Navidad australianas (familia Blandfordiaceae), tubo, garganta y lóbulos.



.jpg)
Periantos bilabiados (y construcciones periantoideas bilabiadas): en una flor cuyo eje se orienta más o menos paralelo al suelo (plagiótropo), el perianto es bilabiado cuando pueden diferenciarse los sectores dorsal y ventral ("ventral" es ante el suelo), llamados respectivamente labio superior o dorsal, y labio inferior o ventral, cada labio puede estar formado por un pétalo o todo un sector del perianto u otra estructura petaloidea. Están adaptados a que un insecto aterrice en el labio inferior e ingrese caminando hasta su recompensa en el mismo labio inferior o la base del perianto, de forma que reciba y transfiera el polen típicamente en su lado dorsal (por anteras y estigmas ubicados contra el labio superior), raramente en algún sector desapercibido de su lado ventral.[55] El ejemplo típico es el de la familia de las labiadas (Lamiaceae). El labio o construcción labiada adaxial es el que se origina más cerca al eje del tallo y el más alejado de la bráctea, y coincide con el labio anterior en flores maduras no resupinadas (que no giraron 180º en su desarrollo), y con el labio dorsal o superior cuando se orientan plagiotrópicamente. De la misma forma el labio o construcción labiada abaxial coincide con el labio posterior en flores que no se resupinan al madurar, y el inferior o ventral si plagiótropas. Nótese que "superior" e "inferior" son términos comunes para referirse a los lado dorsal y ventral de una flor con el eje paralelo al suelo, que no deben confundirse con los sectores "superior" e "inferior" del eje floral desde el punto de vista del observador, como cuando se dice que un ovario es súpero (superior en inglés) o ínfero (inferior en inglés). Cuando las flores se resupinan, el lado adaxial se vuelve anterior y es el que se vuelve ventral al orientarse la flor plagiotrópicamente. Las flores bilabiadas son una adaptación a evitar el robo del polen por parte de las abejas, al suspenderlo sobre su lado dorsal; se originaron muchas veces en la historia de la evolución de las angiospermas, y no necesariamente los labios están formados exclusivamente por el perianto, por ejemplo en los lirios del género Iris la flor posee 3 construcciones bilabiadas independientes, cada una formada por una rama estilar petaloidea como labio dorsal y por un tépalo externo como labio ventral, los dos encerrando un único estambre y una región estigmática. (ver más en Westerkamp C, Classen-Bockhoff 2007. Bilabiate flowers, the ultimate response for bees?[55])
- Perianto bilabiado (construcciones periantoideas bilabiadas)
-
 Una bignoniácea, labio dorsal removido. Se observan los 4 estambres y el estilo, de los que la posición dorsal de anteras y estigma indica que el agente de polinización es el sector dorsal del insecto visitante. En la cara interna del labio ventral se observan guías de néctar y pelos.
Una bignoniácea, labio dorsal removido. Se observan los 4 estambres y el estilo, de los que la posición dorsal de anteras y estigma indica que el agente de polinización es el sector dorsal del insecto visitante. En la cara interna del labio ventral se observan guías de néctar y pelos. -
 Iris (en la foto Iris versicolor). 3 construcciones bilabiadas formadas por una rama del estilo petaloidea como labio dorsal y un tépalo externo como labio ventral, que encierran un estambre y el estigma, escondido en la construcción bilabiada hacia el final de la rama estilar. Alternados a las construcciones bilabiadas los 3 tépalos internos. Como en toda la familia de las iridáceas, falta el verticilo de estambres interno.
Iris (en la foto Iris versicolor). 3 construcciones bilabiadas formadas por una rama del estilo petaloidea como labio dorsal y un tépalo externo como labio ventral, que encierran un estambre y el estigma, escondido en la construcción bilabiada hacia el final de la rama estilar. Alternados a las construcciones bilabiadas los 3 tépalos internos. Como en toda la familia de las iridáceas, falta el verticilo de estambres interno.


- Un perianto o construcción bilabiado y galeado forma con el labio superior una gálea, es decir es cóncavo visto desde el eje de la flor. La gálea puede provenir de cualquier estructura petaloidea. Las anteras se encuentran en el lado dorsal, protegidas por la gálea, muchas veces el visitante "legítimo" al acceder a la recompensa dispara un mecanismo que le acerca el polen a su lado dorsal.[55]
- Gálea
-
 Perianto galeado. Marcado el labio superior con forma de gálea.
Perianto galeado. Marcado el labio superior con forma de gálea. -
 Perianto bilabiado (con labios superior e inferior, o dorsal y ventral) y galeado (el labio dorsal es una gálea) en Salvia microphylla, una lamiácea.
Perianto bilabiado (con labios superior e inferior, o dorsal y ventral) y galeado (el labio dorsal es una gálea) en Salvia microphylla, una lamiácea. -
 Labio superior en forma de gálea en Aconitum.
Labio superior en forma de gálea en Aconitum. -
 Gálea en la orquídea Serapias.
Gálea en la orquídea Serapias.
_-_parts_hood_(upper_lip)_and_lower_lip.svg)
.jpg)


- Un perianto o construcción bilabiado y personado cierra sus labios superior e inferior en una constricción que encierra en el tubo floral los verticilos internos de la flor, es el labio inferior el que se abre y vuelve hacia el labio superior para generar la constricción, luego de ella el labio superior se continúa hacia arriba, y el labio inferior se continúa hacia abajo. De esta manera se impide completamente el acceso al polen si no se fuerza la entrada.[55] Un ejemplo es el conejito, Antirrhinum.
- Perianto personado
-
 Corte longitudinal de labios dorsal y ventral en un perianto personado.
Corte longitudinal de labios dorsal y ventral en un perianto personado. -
 El labio ventral se cierra contra el labio dorsal formando una constricción antes de separarse en el perianto personado del conejito, Antirrhinum.
El labio ventral se cierra contra el labio dorsal formando una constricción antes de separarse en el perianto personado del conejito, Antirrhinum.




- En cambio es unilabiado si posee un solo "labio", como un lóbulo largo con forma de lengua, como en Anigozanthos y en las especies de Aristolochia unilabiadas. Comparar con "labelo".
- Un labelo (no labio) es una pieza o lóbulo del perianto modificado en una estructura compleja y diferente del resto del perianto, típicamente es una pieza más grande que el resto del perianto, y esperablemente gana en complejidad gracias a una anatomía muy vascularizada en la que parece haber piezas fusionadas. El término se utiliza en familias como las orquídeas, que poseen un labelo ancestralmente ventral y preparado para el aterrizaje de insectos, las parasíticas Corsiaceae, cuyo labelo es dorsal, y las zingiberáceas. Véase también "ligulado", que se utiliza en asteráceas, en #Glosario de Asteraceae.
Otros tipos y partes incluyen los siguientes:
- Campanulado, con forma de campana, con un tubo basal acampanado y con lóbulos, como en Campanula. El término también puede ser usado para sépalos o pétalos de piezas libres, que formen una campana.
- Perianto campanulado
-
 Corola campanulada.
Corola campanulada. -
 Corola campanulada en Campanula, la campanita.
Corola campanulada en Campanula, la campanita. -
 Corola campanulada en Clematis hirsutissima.
Corola campanulada en Clematis hirsutissima.



- Urceolado, con forma de urna, expandido en la base y comprimido en el ápice, como en muchas Ericaceae.
- Perianto urceolado
-
 Corola urceolada.
Corola urceolada. -
 Corola urceolada en Andromeda polifolia, una ericácea.
Corola urceolada en Andromeda polifolia, una ericácea.


- Infundibuliforme, en forma de embudo o trompeta, atrompetado, con una base tubular pero que va expandiendo el diámetro a medida que se acerca al ápice, como en Ipomoea.
- Perianto infundibuliforme
-
 Perianto infundibuliforme (atrompetado, en forma de embudo)
Perianto infundibuliforme (atrompetado, en forma de embudo) -
 Perianto infundibuliforme gamopétalo (con los pétalos conados, fusionados entre ellos) en Ipomoea.
Perianto infundibuliforme gamopétalo (con los pétalos conados, fusionados entre ellos) en Ipomoea. -
 Perianto infundibuliforme dialipétalo (con los pétalos sin fusionar) en Amaryllis belladonna.
Perianto infundibuliforme dialipétalo (con los pétalos sin fusionar) en Amaryllis belladonna.



- Tubular, principalmente cilíndrico, como un tubo.
- Rotado, con un tubo corto y miembros amplios orientados a ángulos rectos del tubo, como en Phlox (Flora of Australia menciona a Solanum, el género de la papa, el tomate y la berenjena, como un género que en general presenta flores rotadas).
- Hipocrateriforme ("salverform", "con forma de bandeja"), la base es tubular, con un tubo largo y esbelto, y termina en lóbulos acampanados a ángulos rectos del tubo, como en Plumbago.
- Perianto tubular
-
 Perianto tubular.
Perianto tubular. -
 Perianto tubular en Collomia.
Perianto tubular en Collomia.


- Perianto rotado
-
 Perianto rotado.
Perianto rotado. -
 Perianto rotado en Solanum nigrum.
Perianto rotado en Solanum nigrum.


- Perianto hipocrateriforme
-
 Perianto hipocrateriforme.
Perianto hipocrateriforme. -
 Perianto hipocrateriforme en Plumbago.
Perianto hipocrateriforme en Plumbago.

_in_Hyderabad,_AP_W2_IMG_2437.jpg)
- Saccado (saccate), con una evaginación ventral como un saco o bolsillo.
- Espoloneado (spurred), con espolón (spur).
Los dos muchas veces funcionan como cámaras que almacenan néctar y muchas veces el mismo saco o espolón es nectarífero, posee las glándulas productoras de néctar. (ver en Weberling 1989[51])
- Saco
-
 Perianto saccado, con un saco o bolsillo ventral.
Perianto saccado, con un saco o bolsillo ventral. -
 Sépalo inferior saccado (con un saco o bolsillo) en Impatiens ecalcarata.
Sépalo inferior saccado (con un saco o bolsillo) en Impatiens ecalcarata.

.jpg)
- Espolón
-
 Corola con espolón nectarífero.
Corola con espolón nectarífero. -
 Espolón petaloideo en Linaria.
Espolón petaloideo en Linaria. -
 Espolón petaloideo en Utricularia sandersonii.
Espolón petaloideo en Utricularia sandersonii. -
 Espolones petaloideos en Aquilegia formosa.
Espolones petaloideos en Aquilegia formosa.




-
 Cáliz con espolón nectarífero.
Cáliz con espolón nectarífero. -
 Espolón sepaloideo en Lupinus arbustus.
Espolón sepaloideo en Lupinus arbustus. -
 Espolón nectarífero sepaloideo de la flor de Tropaeolum majus.
Espolón nectarífero sepaloideo de la flor de Tropaeolum majus.

.jpg)

- Carinado o aquillado, con carena o quilla, es decir con un doblez agudo longitudinal (en la línea próximo-distal) que le da aspecto de bote, usualmente ubicado ventralmente en una flor orientada más o menos paralela al suelo (plagiótropa), como en algunas Polygalaceae. La quilla es otra adaptación para evitar el robo de polen por parte de las abejas visitantes, al dificultarles el acceso a las anteras escondidas en el doblez.[56]
- Carena o quilla
-
 Perianto aquillado o carinado, en este caso la corola gamopétala es aquillada, con forma de quilla, la quilla es ventral.
Perianto aquillado o carinado, en este caso la corola gamopétala es aquillada, con forma de quilla, la quilla es ventral. -
 Quilla dorsal en una especie de Erythrina.
Quilla dorsal en una especie de Erythrina. -
 Polygala x dalmaisiana, pétalo inferior aquillado y fimbriado (con flecos, como una orla).
Polygala x dalmaisiana, pétalo inferior aquillado y fimbriado (con flecos, como una orla).



.jpg)
- Caliptrado u operculado, con cáliz y corola fusionados en una gorra que encierra el resto de la flor (caliptra u opérculo), que al madurar la flor cae como una unidad exponiendo el resto de las piezas florales, como en Eucaliptus. El opérculo está formado por los sépalos, los pétalos o los dos.
Otras partes:
- Las guías de néctar desvían la atención de los polinizadores del polen hacia otra recompensa. Son encontradas en las flores polinizadas por insectos, que aterrizan en la pieza que posee las guías de néctar y caminan en la dirección indicada.
- ""Barba" (beard en inglés) se refiere a una línea o zona de tricomas en un perianto o parte del perianto.
- Guías de néctar, barba
-
 Guías de néctar en Calochortus (Liliaceae).
Guías de néctar en Calochortus (Liliaceae). -
 Guías de néctar en Alstroemeria (Alstroemeriaceae).
Guías de néctar en Alstroemeria (Alstroemeriaceae). -
 Iris 'Amethyst Flame'. Guías de néctar y "barba" ("beard").
Iris 'Amethyst Flame'. Guías de néctar y "barba" ("beard").


.jpg)
- Finalmente, la función de atracción visual la pueden cumplir estructuras no petaloideas como los filamentos coloreados de los estambres. Ejemplos son mirtáceas como Callistemon rigidus y el mismo Eucaliptus, y leguminosas mimosóideas como el aromito (Acacia caven).
- Filamentos con función de atracción visual
-
 Filamentos amarillos en Acacia caven, el "aromito".
Filamentos amarillos en Acacia caven, el "aromito". -
 Filamentos colorados en Callistemon rigidus.
Filamentos colorados en Callistemon rigidus.

_IMG_1696.JPG)
- Ver perianto de compuestas (disco y rayo, papus) en #Glosario de Asteraceae al final de este artículo.
- Ver las lodículas de Poaceae en el #Glosario de Poacaeae al final de este artículo.
- Ver el perianto papilionáceo en el #Glosario de Fabaceae al final de este artículo.
- Otras definiciones en relación al perianto
Arreglo del perianto, ciclos, merosidad. Se definió "biseriado", "diclamídeo", "ciclos", "dicíclico", "uniseriado", "triseriado", "tricíclico", "tetraseriado", "tetracíclico", "multiseriado", "aclamídeo", "aperiantado", "flor desnuda", "diclamídeo", "homoclamídeo", "monoclamídeo", "apétalo", "asépalo", "isómero", "merosidad", "anisómero", "bímero", "trímero", "tetrámero", "pentámero" en el artículo Terminología descriptiva de las flores (Botánica).
Fusión del perianto. Se definió "dialisépalo", "dialipétalo", "dialitépalo", "aposepalous", "apopetalous", "apotepalous", "gamosépalo", "gamopétalo", "gamotépalo", "sinsepalous", "sinpetalous", "sintepalous" en el artículo Terminología descriptiva de las flores (Botánica).
Prefloración del perianto. Se definió "prefloración" o "vernación" o "aestivation", "imbricada", "imbricada-alternada", "convoluta" o "contorta", "quincuncial", "valvar", "involuta", en Terminología descriptiva de las flores (Botánica). Ver prefloración vexilar en #Glosario de Fabaceae al final de este artículo.
Androceo
editar- Artículo introductorio: Estambre
"Estambres", "estaminodios", "androceo", "granos de polen", "microsporangio", "anteras", "tecas", "tejido conectivo", "filamento", "dehiscencia de las anteras", ya definidos en la #Introducción a los órganos de la planta y en #Flor.
En el estambre laminar la antera típicamente está en la superficie adaxial de la lámina. En el estambre filamentoso el filamento en general es "filiforme" (cilíndrico y largo), pudiendo poseer otras morfologías. El sésil no tiene filamento, subsésil tiene un filamento corto y rudimentario. El estaminodio puede parecerse tanto a los estambres fértiles que sólo se determina que son estaminodios cuando se busca si han liberado polen fértil, como sucede en las flores femeninas de las sapindáceas, que poseen estambres formadores de polen estéril que nunca se libera. Los estaminodios pueden tener o no un anterodio, una estructura como una antera estéril. Las anteras pueden ser ditecas o monotecas ("una teca") como en las canáceas (achiras).
Los estambres pueden estar provistos de apéndices, pueden presentar tricomas (pelos), o nectarios que secretan néctar, o en general vestitura y color cuya terminología se encuentra en la sección de #Terminología general.
-
 Estambres filamentosos en Amaryllis, ya ocurrió la dehiscencia de la antera y el polen cubre su exterior.
Estambres filamentosos en Amaryllis, ya ocurrió la dehiscencia de la antera y el polen cubre su exterior. -

-
 Estambres "algo laminares" en la flor masculina de Amborella trichopoda, la "angiosperma basal".
Estambres "algo laminares" en la flor masculina de Amborella trichopoda, la "angiosperma basal".


-2.jpg)
- Inserción de los estambres
Por inserción de los estambres puede hacerse referencia a dos cosas diferentes. Puede referirse a si los estambres están ocultos en el perianto, esto es, si la longitud de los estambres excede al perianto o no, son llamados exertos si lo hacen (Caesalpinia pulcherrima, Dolichandra cynanchoides), e inclusos o insertos si no lo hacen y por lo tanto quedan cubiertos por el perianto (Tulipa, Agave).
- Estambres exertos e inclusos
-
 Estambres exertos.
Estambres exertos. -
 En las poáceas se exertan los estambres por fuera de las brácteas protectoras de la flor cuando están listos para la polinización.
En las poáceas se exertan los estambres por fuera de las brácteas protectoras de la flor cuando están listos para la polinización. -
 Estambres exertos en Caesalpinia pulcherrima.
Estambres exertos en Caesalpinia pulcherrima.


_W_IMG_1587.jpg)
-
 Estambres inclusos (insertos).
Estambres inclusos (insertos). -
 ?Estambres inclusos en Tulipa.
?Estambres inclusos en Tulipa.


Los estambres siempre están insertos en el receptáculo, pero por "inserción" también se puede hacer referencia a en qué sector a lo largo de su longitud se separa de la corola un estambre adnato (fusionado) al pétalo. Por ejemplo se puede leer "los estambres están insertos a mitad de camino del tubo de la corola". Los estambres epipétalos o petalostémonos están adnatos a los pétalos ("insertos" en ellos), los episépalos, adnatos a los sépalos ("insertos" en ellos).
- Estambres epipétalos y episépalos
-
 Estambre epipétalo.
Estambre epipétalo. -
 Estambres epipétalos en Pulmonaria officinalis.
Estambres epipétalos en Pulmonaria officinalis.


- Arreglo de los estambres, ciclos, número y posición
"Disposición espiralada", "verticilada", "dídimos", "didínamos", "tetradínamos", "uniseriados", "biseriados", "triseriados", "tetraseriados", "diplostémonos", "antesépalo", "antepétalo", "antisépala", "opositisépala", "alternipétala", "alternisépalos", "opositipétalos", "obdiplostémonos", "monandras", "diandras", "poliandras", "isostémona", "anisostémona", "polistémona" en Terminología descriptiva de las flores#Estambres y en #Terminología general.
- Fusión de los estambres
Los términos vistos en #Terminología general son "distinct" (sin fusionar entre ellos), conados (fusionados entre ellos[57]), libres (sin fusionar a una estructura diferente, aunque pueden estar o no fusionados entre ellos), y adnatos (fusionados a una estructura diferente) pueden ser usados.
Términos más especializados y comunes incluyen apostémonos (sin fusionar ni entre ellos ni a otra estructura), diadelfos, con dos grupos de estambres, cada uno conado solo en sus filamentos, como en muchas Faboideae (Fabaceae), que típicamente tienen 9 estambres fusionados en la mayor parte de su filamento y uno fusionado solo en la base o no fusionado en absoluto,monadelfos, con el grupo de estambres conados por sus filamentos, típicamente en Malvaceae sensu stricto y Bombacaceae (hoy Malvoideae y Bombacoideae en un Malvaceae sensu lato), y sinantéreos, con las anteras conadas pero los filamentos libres, carácter diagnóstico de Asteraceae y también se observa en algunas cucurbitáceas. Si están fusionados por anteras y filamentos son sinandros, como en el palo borracho (Chorisia speciosa) y el zapallo, Cucurbita. Términos vistos más arriba en "inserción" son "epipétalo" o "petalostémono" y "episépalo".
?Si los estambres están fusionados formando varios cuerpos, se llama androceo poliadelfo, como en Hypericum, en que los estambres están soldados formando cinco grupos.[57]
La fusión de los estambres, como la inserción, funciona típicamente para "presentar" los estambres a los polinizadores.
- Eestambres monadelfos, diadelfos, sinandros
-
 Estambres monadelfos en malváceas (en la foto la rosa china, Hibiscus rosa-sinensis). Las 5 cabezuelas coloradas son los estigmas del gineceo que atravesó el tubo estaminal.
Estambres monadelfos en malváceas (en la foto la rosa china, Hibiscus rosa-sinensis). Las 5 cabezuelas coloradas son los estigmas del gineceo que atravesó el tubo estaminal. -
 Sinandro (estambres monadelfos y sinantéreos) en el palo borracho, Chorisia speciosa, otra malvácea.
Sinandro (estambres monadelfos y sinantéreos) en el palo borracho, Chorisia speciosa, otra malvácea. -
 Estambres sinantéreos (ver en #Glosario de Asteraceae al final de este artículo).
Estambres sinantéreos (ver en #Glosario de Asteraceae al final de este artículo).



-
 Estambres diadelfos.
Estambres diadelfos. -
 Estambres diadelfos (abajo, separados del resto de la flor) en una leguminosa (fabácea) papilionóidea. Arriba el gineceo que quedaba semiencerrado por los estambres.
Estambres diadelfos (abajo, separados del resto de la flor) en una leguminosa (fabácea) papilionóidea. Arriba el gineceo que quedaba semiencerrado por los estambres. -
 Una típica flor de leguminosa papilionóidea (Fabaceae Papilionoideae).
Una típica flor de leguminosa papilionóidea (Fabaceae Papilionoideae).

.jpg)
-
 ?Estambres poliadelfos.
?Estambres poliadelfos. -
 ?Estambres poliadelfos en Bombax ceiba.
?Estambres poliadelfos en Bombax ceiba. -
 ?Estambres poliadelfos en Bombax costatum.
?Estambres poliadelfos en Bombax costatum.



- Anteras
- Artículo introductorio: Antera
La inserción de las anteras se refiere a la unión del filamento a la antera. Tipos estándar son basifija, antera unida en su base al ápice del filamento, dorsifija, antera unida dorsalmente y medialmente al ápice del filamento, y subbasifija, antera unida cerca de su base al ápice del filamento. También puede llamarse apicifija[58] a la que se fija al ápice del filamento, es decir es basifija y el estambre se genicula en el punto de inserción de la antera hacia su sector proximal,[1]:293,295 como en muchas Ericaceae[1]:293,295 y en Bignoniaceae.[58] Una antera versátil es la que pivota libremente en el punto de unión con el filamento, las anteras versátiles pueden ser dorsifijas, basifijas o subbasifijas.
-
 Antera basifija.
Antera basifija. -
 Anteras basifijas en Ranunculus repens.
Anteras basifijas en Ranunculus repens. -
 Antera basifija versátil.
Antera basifija versátil. -
 Antera basifija y apicifija en Arbutus, una ericácea.
Antera basifija y apicifija en Arbutus, una ericácea.



-
 Antera dorsifija.
Antera dorsifija. -
 Antera dorsifija versátil.
Antera dorsifija versátil. -
 Antera subbasifija.
Antera subbasifija.



Finalmente, un tipo extremo de antera es el polinio de algunas Orchidaceae y Apocynaceae (ver en la cita[cita 2]).
- Dehiscencia de la antera
"Dehiscencia de la antera" ya fue definida.
- Dehiscencia longitudinal, en que la antera se abre siguiendo una sutura paralela al eje largo de la teca. Otros tipos son raros y específicos de algunos grupos:
- Dehisencia poricida, a través de un poro en el ápice de la teca, como en Ericaceae y Solanaceae. El polen puede atravesar el poro cuando la antera vibra por efecto del viento, hay insectos que pueden extraerlo por sonicación (en inglés buzz pollination), baten los músculos de las alas de forma de hacer vibrar la antera artificialmente.
- Dehiscencia transversal, la sutura corta el eje largo de la antera, como en Lachemilla (Rosaceae) e Hippocratea.[58]
- Dehiscencia valvar, la sutura abre a la antera formando una o más lenguas (valvas), como en Lauraceae.
- Tipos de dehiscencia de la antera
-
 Antera de dehiscencia longitudinal.
Antera de dehiscencia longitudinal. -
 Anteras de dehiscencia longitudinal en Lilium (el polen liberado ya fue transportado).
Anteras de dehiscencia longitudinal en Lilium (el polen liberado ya fue transportado). -
 Antera de dehiscencia poricida.
Antera de dehiscencia poricida. -
 Una ericácea, Chimaphila umbellata, señalados los poros de una antera (violeta), de dehiscencia poricida.
Una ericácea, Chimaphila umbellata, señalados los poros de una antera (violeta), de dehiscencia poricida.

.jpg)

.JPG)
-
 Antera de dehiscencia transversal.
Antera de dehiscencia transversal. -
 Antera de dehiscencia valvar.
Antera de dehiscencia valvar. -
 Laurus nobilis, el laurel, posee una valva en cada teca que se abre de abajo arriba, exponiendo el polen adherido a su cara interna. También se observan los apéndices nectaríferos amarillos a los lados de los filamentos.
Laurus nobilis, el laurel, posee una valva en cada teca que se abre de abajo arriba, exponiendo el polen adherido a su cara interna. También se observan los apéndices nectaríferos amarillos a los lados de los filamentos.


_y_ap%C3%A9ndices_nectar%C3%ADferos.jpg)
Se puede indicar la dirección de la dehiscencia, la dirección hacia la que se expone el polen después de la dehiscencia de la antera, en relación al eje floral o al suelo. La dirección es mejor detectada cuando las anteras todavía están inmaduras o justo empezando a abrirse, porque después de la dehiscencia, las anteras usualmente se resecan y tuercen, volviendo más dificultosa la observación de la dirección de la dehiscencia.
Tipos comunes de dirección de la dehiscencia son:
- extrorsa, que se abre hacia afuera del eje floral,
- introrsa, que se abre hacia adentro del eje floral,
- latrorsa o lateral, que se abre lateralmente al eje floral,
y en flores orientadas horizontalmente, la dehiscencia puede tener dirección
- hacia arriba o
- hacia abajo,
en relación al suelo.
Ejemplos: Xyris tiene una interesante dehiscencia latrorsa o lateral.
La dirección de la dehiscencia puede describirse en dos momentos diferentes del desarrollo. Por ejemplo es un caso común que las anteras sean introrsas temprano en el desarrollo, pero que después con la flor orientada horizontalmente, las anteras se reubiquen en su lado dorsal, con todas sus anteras "mirando" hacia abajo (con la dehiscencia de las anteras hacia abajo). Esta dirección puede ser descripta como introrsa temprano en el desarrollo y hacia abajo en la madurez. Otro ejemplo son las anteras poricidas de Ericaceae, que pueden ser extrorsas temprano en el desarrollo, e introrsas en la madurez por inversión del estambre.
Gineceo, carpelos, pistilos
editar
Si hay más de un ovario, o estilo, o estigma, pero alguno de ellos parece fusionado, todos se consideran parte del mismo pistilo (por ejemplo, 3 ovarios aparentes fusionados en la base, o el caso único de los asclepiadoides, en que el gineceo consiste en dos carpelos que forman distintos ovarios y estilos pero un único estigma uniendo los estilos, como los dos carpelos son conados, toda la estructura es un único pistilo). Un pistilodio[5] es un pistilo estéril, un carpelodio[5] es un carpelo estéril. Los pistilos pueden ser simples (de un solo carpelo) o compuestos por más de un carpelo.
En una sección hecha con un bisturí de hoja delgada y, si la flor es pequeña, a la lupa con ayuda de pinza de punta fina y una aguja de disección, se pueden observar más caracteres, que todavía se consideran dentro del campo de la morfología. Dentro del ovario, un septo es una partición, esperablemente está compuesta por la fusión de las láminas foliares de dos carpelos contiguos. Si los carpelos se encuentran en el centro del eje floral, se llama columna al eje central desde el cual radian los septos, según la disposición de los carpelos puede estar formada por el equivalente al nervio central o el nervio central puede estar en las paredes externas del ovario (la disposición de los nervios en una sección transversal del ovario se considera un carácter anatómico que a veces requiere del uso de un microscopio óptico para su observación). Cada cavidad es un lóculo. Placenta es el tejido de la pared interna del ovario en el que se forma el óvulo, puede haber más de uno en una misma placenta. El funículo es el cordón que une la placenta con el óvulo.
-
 Gineceo de pistilos libres, cada pistilo un carpelo (gineceo dialicarpelar). A la derecha el corte transversal. 3 carpelos y 1 lóculo por carpelo.
Gineceo de pistilos libres, cada pistilo un carpelo (gineceo dialicarpelar). A la derecha el corte transversal. 3 carpelos y 1 lóculo por carpelo. -

-
 Carpelos fusionados en un único pistilo (gineceo gamocarpelar). A la derecha el corte transversal. 3 carpelos y 1 lóculo.
Carpelos fusionados en un único pistilo (gineceo gamocarpelar). A la derecha el corte transversal. 3 carpelos y 1 lóculo. -
 Corte longitudinal con un solo óvulo. 2 carpelos y 1 lóculo.
Corte longitudinal con un solo óvulo. 2 carpelos y 1 lóculo. -
 Gineceo gamocarpelar. A la derecha el corte transversal. 5 carpelos y 5 lóculos, 5 septos y 1 columna.
Gineceo gamocarpelar. A la derecha el corte transversal. 5 carpelos y 5 lóculos, 5 septos y 1 columna. -
 Gineceo unicarpelar. A la derecha el corte transversal. 1 carpelo y 1 lóculo.
Gineceo unicarpelar. A la derecha el corte transversal. 1 carpelo y 1 lóculo.






"Apocárpico" o "dialicarpelar", "sincárpico" o "gamocarpelar", "unilocular", "plurilocular", y cómo determinarlo, en Terminología descriptiva de las flores.
- Placentación
La posición del ovario determina la posición o ubicación del ovario en relación con otras partes florales: al hipanto, al cáliz, a la corola, o al androceo. Un ovario súpero (en flores hipóginas) es el que tiene los sépalos, pétalos, estambres, e hipanto si lo hay, unidos a la base del ovario, como en Prunus y Rosa. Un ovario ínfero (en flores epíginas) es el que tiene los sépalos, pétalos, estambres e hipanto si lo hay, unidos al ápice del ovario, como en Pyrus, Narcissus, Cucurbita. Existe una gama de intermedios entre estas dos condiciones, se puede usar el término semiínfero o semiepígino (o "en parte ínfero") si los sépalos, los pétalos, los estambres y el hipanto si lo hay, están unidos en el medio del ovario, como en Saxifraga, Euonymus y algunos géneros de Celastraceae. Algunos autores[59] hacen descripciones más detalladas y en lugar de "semiínferos" llaman a los ovarios "un quinto ínfero" (solo la quinta parte del ovario ínfera), "un tercio ínfero" (solo la tercera parte del ovario ínfera) etcétera.
El tipo de inserción del ovario y su posición son mejor determinadas en una sección longitudinal de la flor.[5]
- Ovario súpero o ínfero
-
 Ovario súpero. Sin hipanto.
Ovario súpero. Sin hipanto. -
 Ovario súpero en Polygonatum multiflorum. Corte longitudinal de la flor.
Ovario súpero en Polygonatum multiflorum. Corte longitudinal de la flor. -
 Ovario ínfero. Sin hipanto.
Ovario ínfero. Sin hipanto. -
 Ovario semiínfero. Sin hipanto.
Ovario semiínfero. Sin hipanto. -
 Ovario semiínfero, en este caso "un tercio ínfero" (en este caso en relación con los tépalos) en Anticlea elegans.
Ovario semiínfero, en este caso "un tercio ínfero" (en este caso en relación con los tépalos) en Anticlea elegans. -
 Ovario súpero. Con hipanto.
Ovario súpero. Con hipanto. -
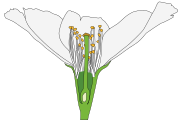 Ovario súpero con hipanto en Prunus sp.
Ovario súpero con hipanto en Prunus sp. -
 Ovario ínfero. Con hipanto.
Ovario ínfero. Con hipanto. -
 Ovario ínfero con hipanto en el zucchini, de la especie Cucurbita pepo. En cucurbitáceas típicamente el ovario se describe como ínfero con hipanto; estando los estambres insertos a diferentes alturas del tubo perigonial según el género.
Ovario ínfero con hipanto en el zucchini, de la especie Cucurbita pepo. En cucurbitáceas típicamente el ovario se describe como ínfero con hipanto; estando los estambres insertos a diferentes alturas del tubo perigonial según el género.









La definición de hipanto varía entre autores. Lo que en Simpson (2005) es un ovario ínfero sin hipanto (ver imagen), según otros autores posee un hipanto que está fusionado al pistilo en toda su extensión.[5] Lo que en Simpson (2005) es un ovario súpero con hipanto, para otros autores es un ovario perígino.[60] En este caso en especial hay que chequear el glosario de la flora que se esté utilizando con cuidado para corroborar cuál es el uso que se le da a estos términos.
Ovario "hipógino", ovario "epígino", "epihipógino", "perígino", "epiperígino", "epihipoperígino" en Terminología descriptiva de las flores (Botánica).
"Placentación axilar", "apical" o "péndula", "apical-axilar", "basal", "libre-central", "laminar", "parietal difusa", "marginal", "parietal", "parietal-septada", "axilar con placentas intrusivas", "axilar con doble curvatura de carpelos" en Terminología descriptiva de las flores (Botánica).
- Estilo
Definido en la #Introducción a los órganos de las plantas
El estilo es de longitud variable, desde menos de 0,5 mm (estigma subsésil) hasta más de 30 cm en ciertas variedades de maíz: es lo que es conoce como "barbas del choclo". Puede ser geniculado como en Gloriosa superba; petaloideo (formando lámina vistosa atractora de polinizadores) como en Canna.,[61] formando una estructura 5-partida similar a una sombrilla, en Sarracenia.[61] El azafrán son los estilos y estigmas secos de Crocus sativus, una iridácea.
La posición del estilo es la ubicación del estilo en relación al cuerpo del ovario. Una posición del estilo terminal o apical es que se origina en el ápice del ovario, y es por lejos el tipo más común. Un estilo subapical se origina de un lado del ovario, cerca del ápice y ligeramente por debajo de él. Un estilo lateral se origina en el lado del ovario, como en algunos miembros de Rosaceae como Fragaria (la frutilla o fresa). Finalmente, un estilo ginobásico es el que se origina en la base del ovario, característicos de Boraginaceae sensu stricto y de la mayoría de Lamiaceae, en que el estilo se eleva desde la base y centro de un ovario fuertemente lobado.
- Estilo terminal, lateral, ginobásico
-
 Posición terminal del estilo.
Posición terminal del estilo. -
 ?Estilo terminal geniculado.
?Estilo terminal geniculado. -
 ?Estilo terminal geniculado en Gloriosa superba.
?Estilo terminal geniculado en Gloriosa superba. -
 Posición lateral del estilo.
Posición lateral del estilo.




-
 El estilo es de posición lateral en Fragaria, la frutilla o fresa.
El estilo es de posición lateral en Fragaria, la frutilla o fresa. -
 Posición ginobásica del estilo.
Posición ginobásica del estilo. -
 El estilo es de posición ginobásica en las boragináceas, en que el ovario se divide en 4 lóbulos que seguirán siendo diferenciables en su fruto.
El estilo es de posición ginobásica en las boragináceas, en que el ovario se divide en 4 lóbulos que seguirán siendo diferenciables en su fruto.


- Morfologías
-
 Las iridáceas perdieron un verticilo de estambres, pero en muchas de ellas, como en Iris (ya vista), las ramas estilares mejoran la eficiencia de los estambres que quedan cumpliendo una función significativa en el síndrome floral, muchas veces son alargadas y petaloideas. El azafrán son los estilos-estigmas curados (secados) de Crocus sativus (en rojizo, 3 ramas, entre las 3 anteras), una iridácea.
Las iridáceas perdieron un verticilo de estambres, pero en muchas de ellas, como en Iris (ya vista), las ramas estilares mejoran la eficiencia de los estambres que quedan cumpliendo una función significativa en el síndrome floral, muchas veces son alargadas y petaloideas. El azafrán son los estilos-estigmas curados (secados) de Crocus sativus (en rojizo, 3 ramas, entre las 3 anteras), una iridácea. -
 El estilo de la carnívora Sarracenia tiene la forma de una sombrilla de 5 puntas (?y cuál es el síndrome floral? no debería haber nectarios, o comen polen? si comen polen dónde se les pega para transporte, en el lado dorsal o el ventral?).
El estilo de la carnívora Sarracenia tiene la forma de una sombrilla de 5 puntas (?y cuál es el síndrome floral? no debería haber nectarios, o comen polen? si comen polen dónde se les pega para transporte, en el lado dorsal o el ventral?).


"Beak" estilar y otras morfologías en Terminología descriptiva de las flores (Botánica).
- Estigma
Definido en la #Introducción a los órganos de la planta
Morfologías (húmedos, secos, papilosos, no papilosos, plumosos, globosos, en cabezuela, capitados, lineal, lobado) en Terminología descriptiva de las flores (Botánica).
- Estigmas
-
 Rosa china, Hibiscus rosa-sinensis, posee 5 estigmas globosos, en cabezuela, capitados.
Rosa china, Hibiscus rosa-sinensis, posee 5 estigmas globosos, en cabezuela, capitados. -
 Zapallo de la especie Cucurbita pepo, se observan 3 estilos blancos (uno removido) que evidencian el número de carpelos, cada uno con un estigma amarillo bilobado.
Zapallo de la especie Cucurbita pepo, se observan 3 estilos blancos (uno removido) que evidencian el número de carpelos, cada uno con un estigma amarillo bilobado. -
 En la parte superior de la foto se observan los pistilos de la flor de Magnolia y sus regiones estigmáticas lineales, de marrón más oscuro (debajo de ellos los estambres, blancos, y luego los pétalos).
En la parte superior de la foto se observan los pistilos de la flor de Magnolia y sus regiones estigmáticas lineales, de marrón más oscuro (debajo de ellos los estambres, blancos, y luego los pétalos). -
 Flores en antesis de Holcus mollis (poáceas, la familia de los pastos y cereales), se observan los estigmas plumosos y los estambres.
Flores en antesis de Holcus mollis (poáceas, la familia de los pastos y cereales), se observan los estigmas plumosos y los estambres.

_secci%C3%B3n_de_perianto_y_ovario_y_1_estilo_removido,_separando_los_dos_estilos_restantes,_etiquetas.jpg)


Asteraceae. Estilo con anillo colector, cepillo en #Glosario de Asteraceae
- Óvulo
- Artículo introductorio: Óvulo
Fueron definidos en la #Introducción a los órganos de la planta y en este artículo se mencionan en #Semilla, ya que algunas partes de la semilla se explican por su desarrollo a partir de partes del óvulo. Otros detalles, probablemente más del campo de la anatomía que de la morfología, ampliados en Terminología descriptiva de las flores (Botánica). En ese artículo: "primordio seminal", "rudimento seminal", "nucela", "tegumentos", "calaza" o "chalaza", "funículo", "micrópilo", "hilo cárpico" o "hilo", "ortótropo", "anátropo" y "campilótropo".
Inflorescencias
editar- Artículo introductorio: Inflorescencia
Las inflorescencias muchas veces funcionan para mejorar la reproducción. Pueden mejorar la capacidad de atracción del polinizador. Otras inflorescencias son pseudantos ("falsa flor"), el síndrome floral se aplica no a una flor individual sino a toda la inflorescencia, como en aráceas y asteráceas. Una dificultad en la determinación del tipo y partes de inflorescencia es delimitarla. Generalmente, una inflorescencia es limitada por la hoja vegetativa más cercana. Sin embargo puede haber una gradación entre las hojas vegetativas más bajas y las brácteas florales pequeñas, por lo que la delimitación de la inflorescencia puede terminar siendo un poco arbitraria. Nótese que la inflorescencia puede consistir en una sola flor por reducción del resto.
La estructura de una inflorescencia puede ser complicada. Ver Weberling (1989[51], al principio no era un fake) para un estudio detallado de las inflorescencias.
"Inflorescencia" y "pedúnculo" fueron definidos en la #Introducción a los órganos de la planta, una inflorescencia con pedúnculo es "pedunculada". Una planta "acaulescente" y "escapo" en #Hábito. En #Brácteas de la flor e inflorescencia se ha visto "involucro" y "espata".
Un raquis es el eje central dentro de la inflorescencia, es la continuación del pedúnculo una vez comienzan las flores o la primera bráctea que se considera parte de la inflorescencia, como la espata en aráceas o la gluma en poáceas (por excepción en poáceas y ciperáceas, cuya inflorescencia se llama "espiguilla", no tienen raquis sino "raquilla").
- Posición de la inflorescencia
Ya se ha visto en la #Introducción a los órganos de la planta que una inflorescencia, que es un módulo, puede ser "axilar", si no es "terminal", en Tallo que la planta es "cauliflora" si la flor o inflorescencia es adventicia en un tronco con crecimiento secundario, y que los fenómenos de "concaulescencia" y "recaulescencia" pueden enmascarar estas posiciones.
"Infrafoliar"; "interfoliar" y "suprafoliar" en #Glosario de Arecaceae al final de este artículo.
- Desarrollo de la inflorescencia
La inflorescencia puede ser, como se ha visto en tallos, determinada ("cerrada", "monotélica") o indeterminada ("abierta" o "politélica"). Característicamente en la inflorescencia indeterminada las flores maduran de la base hacia el ápice, en las determinadas madura el ápice primero.
- Inflorescencias determinadas e indeterminadas
-

-
 ...En cambio en Agave la inflorescencia es determinada.
...En cambio en Agave la inflorescencia es determinada.


También se vio que en algunos casos el ápice de la inflorescencia puede reanudar el crecimiento vegetativo (es un tallo pleonántico), volviendo a formar nomofilos, como ocurre en muchas Myrtaceae y en el ananá (Ananas comosus).
- Tallo pleonántico
-
 ...Los dos ejemplos anteriores son hapaxánticos. La inflorescencia del ananá es pleonántica: luego de dar las flores laterales continúa su crecimiento vegetativo.
...Los dos ejemplos anteriores son hapaxánticos. La inflorescencia del ananá es pleonántica: luego de dar las flores laterales continúa su crecimiento vegetativo.

Una misma inflorescencia a su vez puede estar "compuesta" por unidades de inflorescencias, no necesariamente del mismo tipo. Cuando es así se la llama inflorescencia compuesta (si no, es "simple"). Se las numera por número de orden, como a los tallos, pero al revés que a éstos: la última unidad de inflorescencia (la que tiene las flores directamente insertas en el pedúnculo) es la "inflorescencia primaria", las inflorescencias primarias están insertas en el pedúnculo de la "inflorescencia secundaria", etc. Por ejemplo en pastos (Poaceae), la "inflorescencia primaria" siempre es una espiguilla (una espiga corta que se reconoce porque en su base se encuentran la gluma y la glumela), y lo más común es que estén ordenadas en una inflorescencia secundaria y hasta una terciaria. Ejemplos de cómo se nombran las inflorescencias secundarias son "una panícula de espiguillas", o "un corimbo de cabezas", o "un racimo de espigas".
- Inflorescencias compuestas
-
 En gramíneas (#Glosario de Poaceae) la inflorescencia primaria, en cuyo pedúnculo están directamente insertas las flores, está protegida por dos brácteas (la gluma y la glumela). En el género Hordeum (foto) el pedúnculo tomado por los dedos es el de la inflorescencia secundaria, no tiene gluma y glumela, las inflorescencias primarias (las espiguillas) son las que están insertas en el pedúnculo de la secundaria. Esta inflorescencia es una espiga de espiguillas.
En gramíneas (#Glosario de Poaceae) la inflorescencia primaria, en cuyo pedúnculo están directamente insertas las flores, está protegida por dos brácteas (la gluma y la glumela). En el género Hordeum (foto) el pedúnculo tomado por los dedos es el de la inflorescencia secundaria, no tiene gluma y glumela, las inflorescencias primarias (las espiguillas) son las que están insertas en el pedúnculo de la secundaria. Esta inflorescencia es una espiga de espiguillas. -
 En cambio en este género (Chasmanthium) el pedúnculo es el de la inflorescencia primaria, como indican la gluma y la glumela en la base del raquis. Esta inflorescencia es la espiguilla.
En cambio en este género (Chasmanthium) el pedúnculo es el de la inflorescencia primaria, como indican la gluma y la glumela en la base del raquis. Esta inflorescencia es la espiguilla.
_(cropped).jpg)

- Tipos de inflorescencia
Los términos generales son cimas para las inflorescencias determinadas; para las inflorescencias indeterminadas, las más comunes son el racimo (flores pediceladas), la espiga (flores sin pedicelo), la panícula (con pedicelo, laxamente ramificada, con aspecto de paraguas a medio abrir, las flores más jóvenes en el ápice).
- Inflorescencias indeterminadas
-
 Espiga.
Espiga. -
 Racimo.
Racimo. -
 Racimo.
Racimo. -
 Panícula.
Panícula.




Puede ser determinada o indeterminada la umbela ("como un paraguas", un solo verticilo, las flores a la misma altura en relación al pedúnculo, o a la misma distancia de la inserción al pedúnculo), un radio es el eje secundario de la umbela compuesta (una umbela de umbelas, como en muchas umbelíferas). La cabeza o capítulo es un grupo apretujado de flores sésiles o subsésiles en un "receptáculo compuesto" o torus proveniente del pedúnculo, muchas veces teniendo debajo de ella un involucro de brácteas llamadas filarios con función sepaloide, son típicas de Asteraceae ("las compuestas", denominación antigua por "flores compuestas por flores") y algunos otros grupos como Eriocaulaceae, "las compuestas de las monocotiledóneas", nótese que algunas inflorescencias parecen una "cabeza" pero no tienen receptáculo compuesto. En un amento, como en Quercus o Salix, el eje principal es blando y péndulo,[62] las flores usualmente son inconspicuas y polinizadas por viento,[5] y pueden tener cualquier tipo de arreglo (ser simples o compuestos, determinados o indeterminados...).[5] Un espádice es una espiga con un eje central engrosado y carnoso, típicamente con flores congestionadas y a veces con una espata que lo protege, como en Araceae, Acorus y Typha.
-
 Umbela simple indeterminada.
Umbela simple indeterminada. -
 Umbela simple determinada.
Umbela simple determinada. -
 Amento.
Amento.


.svg)
-
 Cabeza o capítulo.
Cabeza o capítulo. -
 Cabeza o capítulo, otra forma del receptáculo, con involucro por debajo.
Cabeza o capítulo, otra forma del receptáculo, con involucro por debajo. -
 Espádice.
Espádice.



Un pseudanto sensu Faegri y van der Pijl (1979[63]) es una unidad que parece una sola flor, pero típicamente consiste en dos o más flores que funcionan como un único síndrome floral (compárese con euanto, "flor verdadera", y meranto, cada parte en que se divide una flor que funciona como un síndrome floral, como en Iris). Los pseudantos son cabezas (p.ej. Asteraceae) o espádices (p.ej. Araceae).
Los términos "paraclado", "co-florescencia", "cima" o "tirso determinado", "dicasio", "dicasio simple o címula", "dicasio compuesto", "cima compuesta", "cima paniculoide", "panícula de Weberling", "cima racimoide", "cima elongada", "monocasio", "cima helicoide", bóstryx", "cima escorpioide", "cincino", "drepanio", "ripidio", "flabelo", "corimbo", "fascículo", "glomérulo", "pleiocasio", "tirso", "verticiláster", en Terminología descriptiva de las flores (Botánica).
Frutos
editar- Artículo introductorio: Fruto
El "fruto" y "partes accesorias", "pericarpio", "endocarpio", "mesocarpio", "exocarpio" definidos en la #Introducción a los órganos de la planta. "Clamidocarpo", "induvias", "frutos unispermos", "partenocárpicos" en Terminología descriptiva de las flores y frutos (Botánica). "Monotalámico" y "politalámico" en Terminología descriptiva de las flores y frutos (Botánica).
Hay mucha terminología y discrepancia entre autores acerca de qué enfoque se debería dar a la clasificación de los frutos. Suelen concordar en comenzar la clasificación determinando la unidad del fruto.
La unidad del fruto. Los frutos simples derivan de un único pistilo de una única flor, los frutos agregados o polifrutos, de los pistilos (cada pistilo es una unidad fruto) de una sola flor en un gineceo con más de un pistilo, y los frutos múltiples o infrutescencias derivan de más de una flor, cada pistilo es una unidad fruto. El esquizocarpo es un fruto seco derivado de un pistilo en que cada carpelo o lóculo se separa del resto a la madurez, constituyendo cada unidad un fruto en sí mismo (un mericarpio?). Se determina si una planta tiene frutos simples, polifrutos, frutos múltiples o esquizocarpos observando el desarrollo del mismo desde el momento en que las flores son fecundadas. Los frutos individuales se definen de la misma manera que los frutos simples. Los polifrutos son "un fruto agregado de...", las infrutescencias "un fruto múltiple de..." y los últimos son "esquizocarpos de..." la unidad de fruto. Nótese que la unidad de fruto y el fruto compuesto difieren de la "unidad de dispersión", las partes externas a la flor que actúan como partes accesorias (sin serlo, ya que en los libros de texto solo las partes florales se llaman partes accesorias) pueden estar incluidas en la unidad de dispersión, como ocurriría con el involucro derivado de brácteas con función sepaloide que protegen el ovario de la flor de ovario ínfera de las bellotas.
Los frutos se clasifican en primer lugar en secos o carnosos, estos últimos comestibles por algún gremio de animales. La unidad fruto y el polifruto o la infrutescencia pueden diferir ya desde este paso de la clasificación, por ejemplo en la frutilla el fruto derivado de cada pistilo es seco, pero el polifruto es carnoso, la parte carnosa es derivada del receptáculo. Los frutos carnosos son en general adaptaciones para la dispersión de las semillas por animales, el pericarpio carnoso siendo la recompensa (con al menos algunas semillas o bien cayéndose o atravesando el sistema digestivo del animal sin dañarse). Si hay solo dos capas, se llaman exocarpio y endocarpio.
En segundo lugar se indica si el fruto al madurar será dehiscente o indehiscente, es decir si se abre espontáneamente o no a la madurez exponiendo las semillas (esto se aplica a los frutos secos, raramente algún fruto carnoso puede ser dehiscente, pero si es indehiscente normalmente no se indica).
El tipo de fruto se puede seguir especificando desde dos enfoques. Puede hacerse alusión a un carácter morfológico que indique una especialización más alta a un síndrome de dispersión (por ejemplo en frutos carnosos puede dársele importancia al "carozo", un endocarpio leñoso -endurecido- que esconde la semilla, o a un exocarpio leñoso, los dos adaptaciones a un gremio más especializado que los consume y dispersa). Términos más anatómicos (como el número de lóculos y semillas, septos, las partes accesorias, etc.) son útiles para diferenciación taxonómica. Por ejemplo la "legumbre" es el fruto de las leguminosas, y el "pepónide" es el fruto de las cucurbitáceas, al madurar las diferentes especies dentro de cada familia tienen diferentes tipos de dispersión, son frutos que se reconocen desde lo anatómico.
Los tipos más generales nombrados son aquenio (seco, indehiscente), cápsula (seco, dehiscente), baya (carnoso) y drupa (carnoso con "carozo", un endocarpio leñoso).
En general los nombres derivan de alguno de estos cuatro tipos de fruto, que los describe más especializadamente: una nuez (en lenguaje técnico, no coloquial) es un aquenio con un pericarpio muy leñoso, de manera que sólo puede romperla un gremio de animales más especializado que los que pueden dispersar el aquenio; una sámara es un aquenio alado; un grano o cariópside es un aquenio con el pericarpio no leñoso y adnato a los teguentos de la semilla, de forma que sin un análisis de los tejidos de los que proviene pareciera ser la semilla sin pericarpio. "Antocarpo", "Cipsela", "Trima", "Pseudodrupa", "Núcula", "Utrículo", en Terminología descriptiva de las flores y frutos (Botánica)#Frutos.
- Frutos secos indehiscentes (de tipo aquenio)
-
 Aquenio, sección longitudinal.
Aquenio, sección longitudinal. -
 Grano o cariópside, sección longitudinal.
Grano o cariópside, sección longitudinal. -
 Nuez, sección longitudinal.
Nuez, sección longitudinal.



-
 Sámara.
Sámara. -
 Sámaras.
Sámaras.


Entre las cápsulas, las líneas de dehiscencia del pixidio delimitan un opérculo (una "tapita"), la cápsula poricida se abre por poros, el folículo tiene una única línea de dehiscencia longitudinal. "Cápsula loculicida", "cápsula septicida", "cápsula placenticida", "cápsula circuncisa" (pixidio), "cápsula septífraga", "dehiscencia sutural simple o ventricida", "dehiscencia loculicida", "dehiscencia transversal" o "dehiscencia operculada", "dehiscencia poricida o foraminal", "dehiscencia biscida" en Terminología descriptiva de las flores y frutos (Botánica)#Frutos. Para el fruto de Fabaceae (leguminosas), ver "legumbre", "vaina", "lomento", "sutura longitudinal doble", en #Glosario de Fabaceae alf final de este artículo. Para el fruto de Brassicaceae, ver "silicua" y "silícula" en el #Glosario de Brassicaceae al final de este artículo.
- Frutos de tipo cápsula
-
 Inflorescencia de cápsulas aún cerradas en Liquidambar styraciflua.
Inflorescencia de cápsulas aún cerradas en Liquidambar styraciflua. -
 Infrutescencia de cápsulas ya abiertas por sus líneas de dehiscencia en Liquidambar styraciflua.
Infrutescencia de cápsulas ya abiertas por sus líneas de dehiscencia en Liquidambar styraciflua. -
 Folículo en Brachychiton.
Folículo en Brachychiton.


.JPG)
-
 Cápsula circuncisa o pixidio.
Cápsula circuncisa o pixidio. -
 Cápsula circuncisa o pixidio.
Cápsula circuncisa o pixidio.


-
 Cápsula poricida.
Cápsula poricida. -
 Cápsula poricida en la amapola, Papaver. El viento agita el largo pedicelo de manera que las semillas escapan por los poros.
Cápsula poricida en la amapola, Papaver. El viento agita el largo pedicelo de manera que las semillas escapan por los poros.


Frutos carnosos. "Pireno" y "nuculanio", y los especializados de algunos taxones; "hesperidio" o "cítrico" ("flavedo", "albedo", "sacos de jugo"), "pomo", "balaústa" en Terminología descriptiva de las flores y frutos (Botánica)#Frutos. "Pepónide" en #Glosario de Cucurbitaceae al final de este artículo.
Esquizocarpos. "Esquizocarpo de sámaras", ?"esquizocarpo de folículos", ?"esquizocarpo de núculas", "esquizocarpo de aquenios", "regma", en Terminología descriptiva de las flores y frutos (Botánica)#Frutos. "Esquizocarpo de mericarpios" y "carpóforo" en el #Glosario de Apiaceae al final de este artículo.
- Tipos de frutos agregados
El tipo de fruto agregado es indicado como "un fruto agregado de..." las unidades del fruto particulares (o en inglés, agregando el sufijo -acetum al término de la unidad del fruto).
Un poliaquenio (aquenecetum) es un fruto agregado de aquenios. Un ejemplo común es Fragaria, la frutilla o fresa (algunos la llaman conocarpo por tener el receptáculo acrescente carnoso y comestible, y el cáliz persistente), otro que también es de las rosáceas es Rosa (algunos la llaman cinorrodon porque los carpelos libres, unispermos, permanecen envueltos por el receptáculo acrescente, carnoso). Otros ejemplos de poliaquenios: Clematis (una ranunculácea), Ranunculus, Alisma.
- Poliaquenio de la frutilla
-
 Los carpelos libres de Fragaria serán la unidad de fruto del poliaquenio llamado frutilla o fresa.
Los carpelos libres de Fragaria serán la unidad de fruto del poliaquenio llamado frutilla o fresa. -
 El cáliz de la flor persistirá en el fruto.
El cáliz de la flor persistirá en el fruto. -
 Poliaquenio de Fragaria vesca, un conocarpo.
Poliaquenio de Fragaria vesca, un conocarpo. -
 Diagrama de poliaquenio de Fragaria, y el recorrido de los haces vasculares en el receptáculo (rojo), inervando cada aquenio.
Diagrama de poliaquenio de Fragaria, y el recorrido de los haces vasculares en el receptáculo (rojo), inervando cada aquenio.



- Poliaquenio de la rosa
-
 Flor de Rosa.
Flor de Rosa. -
 Formación del fruto en Rosa. En amarillo grisáceo los aquenios con estilos largos.
Formación del fruto en Rosa. En amarillo grisáceo los aquenios con estilos largos. -
 Corte de flor de Rosa.
Corte de flor de Rosa. -
 Fruto de Rosa, el escaramujo o cinorrodon.
Fruto de Rosa, el escaramujo o cinorrodon.




- Poliaquenio de Ranunculus
-
 Los carpelos libres de Ranunculus formarán un poliaquenio.
Los carpelos libres de Ranunculus formarán un poliaquenio. -
 Poliaquenio en Ranunculus.
Poliaquenio en Ranunculus.


Una polidrupa o pluridrupa (drupecetum) es un fruto agregado de drupas, como en Rubus, la frambuesa.
- Pluridrupa en la frambuesa
-
 Los carpelos libres de Rubus formarán la unidad de la pluridrupa.
Los carpelos libres de Rubus formarán la unidad de la pluridrupa. -
 Pluridrupa de Rubus illecebrosus.
Pluridrupa de Rubus illecebrosus. -
 Pluridrupa de Rubus mostrando el corte transversal de una drupa con su carozo (semilla más endocarpo duro).
Pluridrupa de Rubus mostrando el corte transversal de una drupa con su carozo (semilla más endocarpo duro).

.jpg)

Un plurifolículo (un follicetum) es un fruto agregado de folículos, como en Magnolia y en el anís estrellado, Illicium verum.
- Plurifolículo en magnolias
-
 Flor de Magnolia, los carpelos libres darán el fruto agregado de folículos.
Flor de Magnolia, los carpelos libres darán el fruto agregado de folículos. -
 Comienza la formación del fruto en Magnolia.
Comienza la formación del fruto en Magnolia. -
 Algunos folículos comienzan a abrirse.
Algunos folículos comienzan a abrirse. -
 Folículos abiertos, mostrando las semillas (rojas) algunas de ellas ya perdidas.
Folículos abiertos, mostrando las semillas (rojas) algunas de ellas ya perdidas.




Un sincarpo es un fruto agregado, típicamente de bayas, en que las unidades del fruto se fusionan, como en Annona, una anonácea de la que proviene la chirimoya (el fruto de Annona cherimola).
- Sincarpo en la chirimoya
-
 Flor de Annona, se observan los pétalos (carnosos, gruesos) y sépalos (verdes).
Flor de Annona, se observan los pétalos (carnosos, gruesos) y sépalos (verdes). -
 Flor de Annona. En verde el receptáculo y los sépalos, en amarillo los pétalos, en amarillo oscuro los estambres, en amarillo claro los estilos de los carpelos, en circulitos blanquitos los lóculos de cada carpelo.
Flor de Annona. En verde el receptáculo y los sépalos, en amarillo los pétalos, en amarillo oscuro los estambres, en amarillo claro los estilos de los carpelos, en circulitos blanquitos los lóculos de cada carpelo. -
 Annona crassiflora: Formación del fruto. Se pierden los pétalos y los estambres, y todavía se mantienen los estilos y los sépalos.
Annona crassiflora: Formación del fruto. Se pierden los pétalos y los estambres, y todavía se mantienen los estilos y los sépalos. -
 Fruto maduro, sincarpo de bayas, en Annona cherimola, la chirimoya.
Fruto maduro, sincarpo de bayas, en Annona cherimola, la chirimoya.




- Tipos de frutos múltiples
Un fruto múltiple (o infrutescencia) es el derivado de dos o más flores que coalescen. Al determinarlo, también se debe indicar la unidad del fruto que corresponde a un pistilo o a una flor, el fruto entonces se puede llamar "fruto múltiple de..." la particular unidad del fruto presente. Un ejemplo de aquenio múltiple es el de Platanus (a veces vistos como núculas, o como antocarpos debido a los pelos), el plátano de sombra, otro es el panadero de las asteráceas, uno de drupa múltiple es Morus nigra, la mora, donde la parte carnosa son en realidad los tépalos acrescentes.
- Fruto múltiple de aquenios en el plátano
-
 Fruto múltiple de aquenios en Platanus.
Fruto múltiple de aquenios en Platanus. -
 Algunos aquenios removidos.
Algunos aquenios removidos. -
 Un aquenio.
Un aquenio. -
 Corte transversal mostrando la semilla.
Corte transversal mostrando la semilla.

_2.jpg)

.svg)
- Drupa múltiple en la mora
-
 Inflorescencia de flores femeninas en la mora, Morus nigra.
Inflorescencia de flores femeninas en la mora, Morus nigra. -
 Drupa múltiple en Morus nigra.
Drupa múltiple en Morus nigra.


Algunos términos especializados para frutos múltiples son los que siguen:
Se llama "bur" (abrojo en Argentina) cuando el fruto múltiple de aquenios o granos está rodeado de una estructura dura y espinosa derivada de un involucro, con capacidad de adherirse a los animales que se trasladan, como en Cenchrus (Poaceae) o Xanthium (Asteraceae). El abrojo es una de las estrategias de la ectozoocoria (la dispersión de la semilla por animales sin ingestión).
- Abrojos
-
 Inflorescencias de Xanthium, una compuesta. Cada inflorescencia de flores violetas está rodeada de un involucro verde. Cada flor madurará en un aquenio, y el involucro los envolverá formando el "bur".
Inflorescencias de Xanthium, una compuesta. Cada inflorescencia de flores violetas está rodeada de un involucro verde. Cada flor madurará en un aquenio, y el involucro los envolverá formando el "bur". -
 Formación del "bur" en Xanthium (las inflorescencias terminales más pequeñas son las masculinas).
Formación del "bur" en Xanthium (las inflorescencias terminales más pequeñas son las masculinas). -
 "Bur" maduro y listo para dispersarse.
"Bur" maduro y listo para dispersarse. -
 "Bur" en Xanthium, en marrón oscuro las unidades aquenios, en marrón más claro la estructura de brácteas involucrales.
"Bur" en Xanthium, en marrón oscuro las unidades aquenios, en marrón más claro la estructura de brácteas involucrales.




-
 Inflorescencia en Arctium, otra asterácea, obsérvese el involucro con ganchos (verde).
Inflorescencia en Arctium, otra asterácea, obsérvese el involucro con ganchos (verde). -
 "Bur" en Arctium.
"Bur" en Arctium. -
 Inflorescencia en cierta ciperácea, Cenchrus.
Inflorescencia en cierta ciperácea, Cenchrus. -
 Frutos maduros de Cenchrus encerrados en un "bur" o abrojo.
Frutos maduros de Cenchrus encerrados en un "bur" o abrojo.


.jpg)

Un sorosis es un fruto múltiple en que las unidades son bayas carnosas y están fusionadas lateralmente a lo largo de un eje central, como en Ananas, el ananá o piña.
- Sorosis en el ananá
-
 Planta de Ananas comosus, la piña o ananá.
Planta de Ananas comosus, la piña o ananá. -
 Inflorescencia (en violeta las flores, en rosa las brácteas).
Inflorescencia (en violeta las flores, en rosa las brácteas). -
 Formación del fruto y la infrutescencia.
Formación del fruto y la infrutescencia. -
 Diagrama de infrutescencia y fruto.
Diagrama de infrutescencia y fruto.




Un siconio o sicono es un fruto múltiple en que las unidades aquenios se presentan en la cara interna de un receptáculo convexo, que se vuelve acrescente y carnoso (siendo la infrutescencia derivada de su inflorescencia particular, el hipantodio), como en Ficus, la higuera.
- Sicono en la higuera
-
 Hipantodio justo antes de la antesis (en este caso la apertura del poro, u ostíolo, del receptáculo formando un recipiente, en la foto el ostíolo cerrado en primer plano), en el higo comestible, Ficus carica.
Hipantodio justo antes de la antesis (en este caso la apertura del poro, u ostíolo, del receptáculo formando un recipiente, en la foto el ostíolo cerrado en primer plano), en el higo comestible, Ficus carica. -
 Sicono en Ficus carica, el higo comestible.
Sicono en Ficus carica, el higo comestible.


Una bibaya está formada por la concrescencia de dos bayas, como en Lonicera japonica, la madreselva. Un estróbilo posee el eje de la inflorescencia cilíndrico, seco, y los frutos en la axila de brácteas leñosas, como en Alnus jorullensis, el aliso, o Casuarina stricta, la casuarina.
-
 Inflorescencias de 2 flores en la madreselva, Lonicera japonica.
Inflorescencias de 2 flores en la madreselva, Lonicera japonica. -
 Bibaya en la madreselva, Lonicera japonica.
Bibaya en la madreselva, Lonicera japonica. -
 Estróbilos de Casuarina stricta.
Estróbilos de Casuarina stricta.



Semillas
editar- Artículo introductorio: Semilla
Los aspectos de la morfología de la semilla pueden ser importantes caracteres utilizados en Sistemática. Algunos aspectos valiosos son el tamaño y la forma, el color, y los caracteres de la superficie del tegumento de la semilla o testa o cubierta seminal o episperma, la cubierta protectora más externa de la semilla derivada del o los tegumentos. Una testa que es carnosa a la madurez puede ser llamada sarcotesta (si bien esto puede ser confundido con un arilo, que está separado de los tegumentos de la semilla). También son importantes la forma, tamaño, y color del hilo, una cicatriz de adherencia del funículo al tegumento de la semilla, y del rafe, una cresta en el tegumento de la semilla formado por un funículo adnato (en óulos anátropos y campilótropos). Algunas semillas tienen un arilo (son ariladas), un crecimiento carnoso del funículo, del rafe, o de la testa (pero separado de la testa) que generalmente funciona en la dispersión de la semilla por animales. Los arilos pueden ser característicos de ciertos grupos, como de Sapindaceae. Similar al arilo es el carúnculo, una expansión carnosa en la base de la semilla, los carúnculos también funcionan en la dispersión por animales, como las semillas carunculadas de Viola, las violetas, que poseen semillas dispersadas por hormigas.
Los óvulos son ortótropos, anátropos o campilótropos como se ve en Terminología descriptiva de las flores y frutos (Botánica), y eso explica las partes de la semilla vistas en morfología. Las semillas derivadas de óvulos ortótropos, por ejemplo, carecen de rafe.
- Tipos de óvulo - tipos de semilla
-
 Óvulo ortótropo. 1: saco embrionario (gametofito femenino), rodeado del nucelo (esporangio) y los 2 tegumentos 2: calaza 3: funículo.
Óvulo ortótropo. 1: saco embrionario (gametofito femenino), rodeado del nucelo (esporangio) y los 2 tegumentos 2: calaza 3: funículo. -
 Óvulo anátropo. 1: saco embrionario (gametofito femenino), rodeado del nucelo (esporangio) y los 2 tegumentos 2: calaza 3: funículo 4: rafe.
Óvulo anátropo. 1: saco embrionario (gametofito femenino), rodeado del nucelo (esporangio) y los 2 tegumentos 2: calaza 3: funículo 4: rafe. -
 Óvulo campilótropo. 1: saco embrionario (gametofito femenino), rodeado del nucelo (esporangio) y los 2 tegumentos 2: calaza 3: funículo.
Óvulo campilótropo. 1: saco embrionario (gametofito femenino), rodeado del nucelo (esporangio) y los 2 tegumentos 2: calaza 3: funículo.



-
 Semilla de leguminosa. La cicatriz que queda luego de la absición del funículo se llama hilo.
Semilla de leguminosa. La cicatriz que queda luego de la absición del funículo se llama hilo.

Algunas semillas poseen apéndices atractivos para las hormigas llamados eleosomas o elaiosomas (literalmente "cuerpos grasos"), ricos en aceites (según algunos autores es un arilo rico en aceites). Las hormigas recolectan esas semillas transportándolas hasta sus nidos, donde el elaiosoma es separado activamente y la semilla abandonada. Un ejemplo de elaiosomas lo poseen las semillas de turneráceas. El opérculo es una proliferación del endostoma, es decir del tegumento interno alrededor de la micrópila. Se encuentra especialmente en monocotiledóneas. El estrofíolo es una proliferación glandular o esponjosa que se forma sobre el rafe, como en Chelidonium majus (Papaveraceae).[64]
El micrópilo puede permanecer como un poro ocluido, como ocurre en Cucurbita o Phaseolus, o puede obliterarse totalmente como ocurre en la semilla de Ginkgo. Los tricomas o pelos son propios de semillas pequeñas, contenidos frecuentemente en frutos de dehiscencia lenta, por ejemplo en las semillas de asclepiadáceas. Los pelos de las semillas del algodón pueden medir de 10 a 65 mm, y se conocen comercialmente como "fibra de algodón". También puede haber semillas aladas, como en Luehea donde el funículo se desarrolla en ala.[64]
Detalles específicos del embrión (el esporófito inmaduro en estado de latencia) pueden ser estudiados. Estos incluyen aspectos del epicótilo' (o plúmula o gémula, el vástago inmaduro) o la radícula (la raíz primaria inmadura, no debe ser confundido con una posición "radical"), el hipocótilo (la región de transición entre el vástago y la raíz), y el cotiledón o los cotiledones (las primeras hojas del embrión, muchas veces con la función de almacenamiento). En las gimnospermas el embrión presenta generalmente dos a varios cotiledones. Las dicotiledóneas en general presentan dos cotiledones que pueden tener diverso aspecto, las monocotiledóneas presentan un solo cotiledón. Algunos miembros de Poaceae, la familia de los pastos, tienen el epicótilo rodeado de una vaina protectora llamada coleóptilo, y la radícula rodeada de una vaina protectora llamada coleorriza. La prefoliación del cotiledón (o "ptyxis") puede ser un carácter valioso.
-
 Germinación de una leguminosa (una dicotiledónea). Se observan los dos cotiledones engrosados con función de almacenamiento.
Germinación de una leguminosa (una dicotiledónea). Se observan los dos cotiledones engrosados con función de almacenamiento.

En las pteridófitas el embrión es unipolar: solo se establece el polo de crecimiento correspondiente al vástago, pues las raíces son adventicias. En cambio en las plantas con semilla, el embrión es bipolar: en un polo se desarrolla el vástago, y en el otro la raíz. En Poaceae el embrión es bastante complejo.
Tipo de endosperma de la semilla
editarLa doble fecundación en angiospermas da lugar al desarrollo del embrión por un lado y del endosperma, tejido nutricio, por otro lado. Todas las angiospermas forman endosperma, el tejido que funciona como almacenamiento de energía derivado de la fusión del esperma con los polos nucleares del gametofito femenino. En gimnospermas hay semilla pero en general no hay doble fecundación con formación de endosperma, y el tejido nutricio es almacenado en el protalo o gametofito femenino haploide. La típica semilla de angiosperma es albuminada o endospermada, es decir la reserva energética está contenida en el endosperma de las semillas maduras. En algunas angiospermas el endosperma se desarrolla, pero muy poco o nada es depositado en las semillas maduras, son llamadas semillas exalbuminadas, como en las orquídeas.
-
 Disección de una semilla de Ginkgo (una gimnosperma), se observa el embrión bipolar (con plúmula y las dos primeras hojas, y radícula), el tejido de reserva es proveniente del gametofito haploide. La testa es de sólo 1 tegumento.
Disección de una semilla de Ginkgo (una gimnosperma), se observa el embrión bipolar (con plúmula y las dos primeras hojas, y radícula), el tejido de reserva es proveniente del gametofito haploide. La testa es de sólo 1 tegumento.

Algunas semillas son perispermadas: el perisperma es el tejido nuclear que perdura y se carga de sustancias de reserva, como en Chenopodiaceae, Amaranthaceae, Polygonaceae, y se considera un carácter primitivo. Algunas semillas perispermadas tienen también endosperma, como las de pimienta (Piper nigrum), Nymphaeaceae y Zingiberaceae.
Finalmente algunas angiospermas son cotilespermas, en que la mayor parte de la reserva de alimento está en los cotiledones, por ejemplo en los porotos o frijoles y en los guisantes.
Tipo de germinación de la semilla
editarEsto requiere la observación de las plántulas jóvenes durante la germinación y describe la posición de los cotiledones durante el proceso. Las plantas con germinación hipógea (criptocotilares) tienen cotiledones que se quedan en el suelo durante la germinación. Las plantas con germinación epígea (fanerocotilares) tienen cotiledones que se elevan sobre la superficie del suelo durante la germinación (según Duke, 1969,[65] la germinación es fanerocotilar cuando los cotiledones emergen del episperma, y criptocotilar cuando no lo hacen).
Se llaman semillas vivíparas las que no tienen estado de latencia. El embrión simplemente crece fuera de la semilla y del fruto estando todavía sobre la planta madre, como en Rhizophora mangle, el mangle.[64]
Terminología general
editarMuchos términos pueden aplicarse a un número de órganos diferentes. Aquí se definen.
Color
editarColor es la medida de las ondas electromagnéticas de luz que se reflejan o transmiten a través de un objeto. Cuando se describe el color, se debe definir bien a qué órgano o parte del órgano se está haciendo referencia. Por ejemplo, en lugar de decir "flores amarillas", se debe describir como "corolas y filamentos amarillos, anteras marrones, polen blanco, ovario verde". El color mismo puede ser definido en una forma muy precisa, usando componentes de tono, valor y croma. Para la precisa designación del color se necesita un patrón de colores (ver Tucker et al. 1991[66]).
El patrón de color es una medida de la distribución de colores en un objeto. Términos comunes son maculado, que quiere decir con puntos pequeños en un fondo más o menos uniforme, pelúcido, con puntos o parches translúcidos, y variegado, con dos o más colores en varios patrones irregulares, generalmente usado para las hojas.
En una hoja o un órgano de dos caras, cuando ambas caras son del mismo color, la hoja se llama concolora; cuando son de distinto color, en el caso de la hoja generalmente es la cara adaxial la que es de color verde más oscuro, la hoja se llama discolora.[67]
Tamaño
editarGeneralmente el tamaño se refiere a medidas lineales, como la altura o el ancho. Deben usarse medidas métricas.
Número
editarNúmero se refiere al conteo de partes. En estructuras verticiladas, se debe hacer una distinción entre ciclo y merosidad.
Ciclo se refiere al número de ciclos o verticilos de partes. Puede ser designado con un número, o pueden ser usados términos como monocíclico o uniseriado, y dicíclico o biseriado, etc. El ciclo es más comúnmente usado para designar el número de ciclos del perianto o del androceo.
Por merosidad se entiende el número de piezas que compone cada verticilo. La merosidad puede ser designada como un número, o pueden ser usados términos como bímero, trímero, tetrámero, pentámero, etc. Dos términos que se usan son isómero, que tiene el mismo número de piezas en verticilos diferentes, y anisómero, con diferente número de piezas en verticilos diferentes. La merosidad es más comúnmente utilizada para las partes de la flor, como el cáliz, la corola, el androceo y el gineceo (equivalente al número de carpelos).
Textura
editarLa textura es la consistencia estructural interna de un objeto, algunos términos de textura también tienen en cuenta el color. La textura es muchas veces utilizada para las hojas pero también puede ser usada para cualquier parte de la planta, como las brácteas o las partes de la flor. La textura puede estar correlacionada con el hábitat de la planta y puede ser representativa de la cantidad de agua almacenada en los tejidos, fibras, haces vasculares, lignina, suberina o cualquier otra característica anatómica de la planta. Términos comunes incluyen: cartilaginoso, con la textura del cartílago, duro pero flexible, usualmente blancuzco. Papiráceo, opaco y con la textura del papel para escribir. Coriáceo, grueso y con la textura del cuero, pero flexible. Herbáceo, con una textura suave y ligeramente suculenta. Indurado, duro e inflexible. Membranoso, delgado y algo translúcido, como una membrana. Mesofítico, con una textura intermedia entre coriáceo y membranoso (típico de muchas hojas comunes). Ruminado, con textura irregular, toscamente arrugado, que se ve como si estuviera masticado (como en el endosperma de Annonaceae). Escarioso, delgado y aparentando seco, usualmente blancuzco o amarronado. Suculento o carnoso, carnoso y jugoso. Leñoso, con una textura dura como la madera.
Fusión
editarFusión se refiere a la aparente unión o falta de unión de las piezas u órganos. Los objetos "fusionados" pueden haberse desarrollado por separado y tomar contacto y unirse después. Este proceso se llama fusión congénita y puede ocurrir por ejemplo en la fusión de las anteras. Sin embargo es más común que los objetos aparentemente fusionados en realidad se desarrollen del mismo tejido meristemático. En este proceso, el primordio que normalmente crecería en objetos separados está elevado por tejido basal que está creciendo continuamente, un típico ejemplo es la "fusión" de la corola y de los estambres (epipetalía).
Los términos relacionados con la fusión tienen en cuenta si son entre piezas iguales o distintas. Conado es la fusión de piezas iguales. Adnato es la fusión de piezas diferentes. "Estambres conados" quiere decir que están unidos entre sí, "Estambres adnatos" significa que están unidos a otra cosa (en general a la corola). Dos términos que refieren al grado de fusión, similares, son coherente, con piezas iguales unidas pero solo superficialmente y fácilmente separables, y adherentes, con piezas diferentes unidas pero solo superficialmente y fácilmente separables.
Algunos términos designan la falta de fusión. "Distinct" significa con partes del mismo tipo sin fusionar entre ellas, p.ej. estambres no conados. Libres significa que las partes no están fusionadas a partes de distinto tipo, p.ej. estambres no fusionados a otras piezas florales. Finalmente, contiguo significa con las piezas tocándose pero no conadas ni adnatas ni coherentes ni adherentes. Las piezas contiguas pueden parecer fusionadas, pero solo están en contacto cercano.
Forma
editarLos términos de forma pueden usarse para tallos, hojas, folíolos u otras partes de la hoja, brácteas, sépalos, pétalos, estambres, pistilos, tricomas, u otras partes de la planta. La forma puede clasificarse como sólida (tridimensional) o plana (en dos dimensiones). La forma plana puede dividirse en la forma general del plano, la base, el margen, class=centerel ápice, el proceso apical, y la forma de la división.
Formas tridimensionales
editarMuchos términos específicos son utilizados. Capitado es con forma de cabeza, esférico con un tallito corto. El término para esférico es globoso, para la forma de una semiesfera es hemisférico. Una forma elipsoide con el eje largo paralelo al punto de unión se llama prolado, cuando el eje largo es perpendicular al punto de unión se llama oblado. Clavado significa con forma de garrote, cilíndrico con un final redondeado y engrosado.
-
 Forma tridimensional capitada.
Forma tridimensional capitada. -
 Forma tridimensional prolada.
Forma tridimensional prolada. -
 Forma tridimensional oblada.
Forma tridimensional oblada. -
 Forma tridimensional clavada.
Forma tridimensional clavada.




Discoide es con forma de disco, y fusiforme es con forma de huso, delgadamente elipsoide con dos fines atenuados. Filiforme significa como una hebra y filamentoso, siendo largo, delgado, y típicamente flexible. Fistuloso significa cilíndrico y hueco por dentro.
-
 Forma tridimensional discoide.
Forma tridimensional discoide. -
 Forma tridimensional fusiforme.
Forma tridimensional fusiforme. -
 Forma tridimensional filiforme.
Forma tridimensional filiforme. -
 Forma tridimensional fistulosa.
Forma tridimensional fistulosa.




Lenticular significa con forma de lente, un disco con los dos lados convexos. Ligulado es con forma de lengua, aplanado y de alguna forma oblongo en forma, como en algunas Asteraceae. Pilado significa con un eje largo cilíndrico terminado en un engrosamiento globoso o elipsoide, como en los tricomas glandulares pilados. Terete es el término general para designar cilíndrico. Turbinado significa con forma de turbante, como en las cabezas turbinadas de Asteraceae.
-
 Forma tridimensional lenticular.
Forma tridimensional lenticular. -
 Forma tridimensional ligulada.
Forma tridimensional ligulada. -
 Forma tridimensional pilada.
Forma tridimensional pilada.



-
 Forma tridimensional terete.
Forma tridimensional terete. -
 Forma tridimensional turbinada.
Forma tridimensional turbinada.


Además de estos términos específicos, otros términos tridimensionales pueden ser derivados de términos bidimensionales agregando el sufijo -oide, como en "elipsoide", "ovoide", etc.
Formas bidimensionales
editarLas formas bidimensionales fueron estandarizadas (ver Systematics Association Committee for Descriptive Terminology, 1962[68]). Estos términos de forma están basados, en parte, en la relación entre el tamaño del largo respecto al ancho de la forma.
Los términos estandarizados por la Asociación (1962[68]): "acicular", "ensiforme", "acintado", "linear", "delgadamente oblongo", "oblongo", "delgadamente elíptico", "elíptico", "ampliamente elíptico", "orbicular", "lanceolado", "lanceovado", "ovado", "ampliamente ovado", "muy ampliamente ovado", "oblanceolado", "oblanceovado", "obovado", "ampliamente obovado", "muy ampliamente obovado", "delgadamente triangular", "triangular", "ampliamente triangular", "deltado", "rómbica", "trulada", "cordada", "cordiforme", "obcordada", "falcada" o "falciforme", "lirada", "pandurada", "reniforme", "espatulada", "subulada", en Terminología descriptiva de las hojas (Botánica)#Formas bidimensionales.
Base foliar "atenuada", "delgadamente cuneada", "cuneada", "aguda", "obtusa", "truncada", "redondeada", "cuneada", "reniforme", "auriculada", "hastada", "sagitada", "oblicua", "peltada", "envainador" en Terminología descriptiva de las hojas (Botánica)#Formas bidimensionales.
Ápice de forma bidimensional "acuminado", "caudado", "cuspidado", "delgadamente agudo", "agudo", "obtuso", "truncado", "redondeado", "oblicuo", "emarginado", "retuso", "proceso apical", "apiculado", "aristado", "cirroso", "mucronado", "mucronulado", "espinoso", "pungente", en Terminología descriptiva de las hojas (Botánica)#Formas bidimensionales.
Combinaciones de formas. La forma general, la forma de la base, y la forma del ápice pueden ser usadas en combinación para describir una variedad de formas bidimensionales. Por ejemplo puede haber hojas que tengan todas una forma general más o menos elípticas pero diferan en la forma de la base y el ápice.
Márgenes
editarLos márgenes y dientes se encuentran en las formas bidimensionales, como las hojas.
"Margen", "diente", "margen entero", "margen dentado", "margen serrado", "margen doblemente serrado", "margen biserrado", "margen denticulado", "margen crenado", "margen crenulado", "toscamente..." o "finamente...", "espaciadamente...", "espinoso", "premorso", "ciliado", "ciliolado", "aciliado", "filífero", "diente violoide", "diente cloranthoide", "diente malvoide", "diente rosoide", "diente salicoide", "diente teoide", "diente espinoso", "diente rutoide", en Terminología descriptiva de las hojas (Botánica)#Márgenes.
División
editar"Lobado", "hendido" (cleft), "partido", "sectado", "pinadamente", "palmadamente", "pinatifido", "pinatisecto", "bipinatifido", "palmatifido", "palmatisecto", "descompuesto", "disecto", "bífido", "inciso", "sinuado", "lacerado", "laciniado", "pectinado", en Terminología descriptiva de las hojas (Botánica)#División.
Disposición
editarDisposición se refiere a la posición relativa de los objetos, en este caso por ejemplo de los órganos de la planta o de las piezas de los órganos. La disposición está dividida en posición, arreglo, orientación y postura.
Posición
editarLa posición es la ubicación de las partes una en relación con la otra, partes no iguales. Algunos términos generales de posición que pueden ser aplicados a hojas, brácteas o piezas de la flor, tienen que ver con el desarrollo. Adaxial (también conocido como ventral) se corresponde con la superficie superior o interna de un órgano. Adaxial literalmente significa "hacia el eje", en el desarrollo temprano de las hojas o partes florales, es la superficie que inicialmente está en cara a o cerca del eje la que típicamente será la superficie superior. La confusión entra cuando el órgano se tuerce más tarde en el desarrollo, en estos casos, lo más claro es decir que la superficie del órgano es adaxial en su desarrollo. Abaxial (también conocido como dorsal) se corresponde con la superficie más baja o externa de un órgano, es decir, la superficie más lejana al eje temprano en el desarrollo. Nótese que ventral y dorsal se usa en forma opuesta a como se usa en animales, por esta razón es que estos términos son evitados en las descripciones de plantas.
Con respecto a una estructura orientada horizontalmente, se utiliza posterior para el lóbulo o parte superior, y anterior para el lóbulo o parte inferior. Posterior y anterior son muy usados en piezas florales de flores orientadas horizontalmente y se corresponden respectivamente con adaxial y abaxial. Basal o radical (no confundir con la radícula del embrión) indica en o cerca de la base de la estructura. Proximal es similar a basal o radical y significa cerca del punto de origen o de unión, como el punto de unión de una hoja al tallo. Apical o terminal significa en el tope o cerca de él, de la punta, o del final de una estructura. Distal es similar a apical o terminal y significa lejos del punto de origen o del punto de unión de un órgano y es especialmente valioso para estructuras que se giran de forma que "base" y "ápice" resultan confusos. Otros términos generales son lateral o axilar, en el lado de una estructura o en la axila (la región adaxial de un nudo), central, en o cerca del centro (plano medio) de la estructura, y circunferencial, en o cerca de la circunferencia de una estructura redondeada.
Algunos términos de posición son usados primariamente para describir la posición de estructuras con respecto al tallo. En este sentido, los términos generales radical o basal significan posicionados en la base del tallo, caulinar significa posicionado a lo largo del tallo, y acrocaulis significa posicionado en el ápice del tallo.
Arreglo
editar
Arreglo es la ubicación de partes en relación con otras partes similares. Algunos términos usados primariamente para hojas, brácteas y piezas florales, describen el número de órganos por nudo. Alternado ("disposición alterna") se refiere a una hoja u otra estructura por nudo. Subcategorías de alternado son monístico, alternado con puntos de unión en una línea vertical (en un ortóstico) como en Costaceae, dístico, alterno con los puntos de unión en dos líneas verticales opuestas (en dos ortósticos[69]) como en Poaceae, trístico, aternado con los puntos de unión en tres líneas verticales (en tres ortósticos), como en Cyperaceae, y espiralada (también llamada polística), alternada con los puntos de unión en más de 3 líneas verticales (se dice que se disponen en más de 3 espirósticos, no llamados ortósticos porque no necesariamente son exactamente verticales, el dibujo que formamos en el tallo debido a la unión imaginaria de los nudos en una línea en espiral es la espira generatriz, y mirando desde el ápice, cuando las hojas son numerosas y los entrenudos son muy cortos, como ocurre en las crasuláceas o en las piñas de las coníferas, se observan alineaciones secundarias denominadas parásticos, que marcan las líneas de contacto de cada hoja con las anteriores o sucesivas, el ángulo de divergencia se determina trazando dos líneas horizontales que pasen por el centro del tallo, cada una atravesando el eje de dos hojas sucesivas, y calculando el ángulo que dejan mirando desde el ápice hacia abajo, comúnmente el ángulo de divergencia es constante para todas las hojas de una misma rama[69]).
Opuesta ("disposición opuesta") se refiere a dos hojas u otra estructura por nudo, en lados opuestos del tallo o eje central. Dos subcategorías de opuesto son decusadas, opuestas con las piezas del par siguiente a ángulos rectos del par anterior (quedando en forma de cruz), y no decusadas, opuestas con las estructuras en ángulos no rectos con respecto al par anterior. La mayoría de las hojas opuestas son decusadas, de hecho las hojas no decusadas pueden ser el resultado de una torcedura del tallo. El arreglo decusado es el tipo de arreglo más comúnmente encontrado en plantas vasculares.[5] Los folíolos de una hoja compuesta son típicamente no decusados. El término subopuesto refiere a dos hojas u otras estructuras en lados opuestos del tallo o eje central pero en diferentes nudos ligeramente desplazados uno con respecto al otro. Verticilado significa con 3 o más piezas por nudo.
-
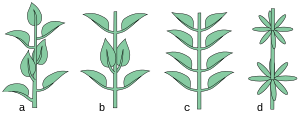 a) Hojas alternas. b) Hojas opuestas decusadas c) Hojas opuestas, en este caso no decusadas. d) Hojas verticiladas
a) Hojas alternas. b) Hojas opuestas decusadas c) Hojas opuestas, en este caso no decusadas. d) Hojas verticiladas -
 Hojas verticiladas.
Hojas verticiladas.


Más términos de arreglo denotan condiciones más especializadas. Equitantes significa hojas con bases que se superponen, usualmente fuertemente dobladas a lo largo de la vena media. Fasciculado significa que las hojas u otras estructuras están en un fascículo, un tallo corto, una agrupación de nudos con entrenudos muy cortos. Imbricado es un término general para estructuras que se superponen. Valvado significa que los lados de dos o más estructuras están girados uno hacia el otro, de forma que se tocan en los márgenes. Rosulado significa en una roseta, un arreglo con partes (usualmente hojas) que irradian de un punto central al nivel del suelo (por ejemplo en Taraxacum officinale, el diente de león). Unilateral generalmente hace referencia a las flores, inflorescencias u otras estructuras en un lado del eje, muchas veces debido a la torcedura de los tallos.
Orientación
editarOrientación hace referencia al ángulo en que se encuentra una estructura con respecto a un eje central muchas veces vertical. Términos precisos de orientación utilizan rangos de ángulos en grados con respecto al eje, en su parte superior, en su parte inferior, o en la horizontal. Los términos son, del más alto al más bajo: oprimido, apretado cercanamente al eje hacia arriba, con un ángulo de divergencia de entre 0º y 15º desde la parte superior del eje, ascendente, direccionado hacia arriba, con un ángulo de divergencia de entre 15º y 45º desde la parte superior del eje, inclinado, direccionado hacia arriba, con un ángulo de divergencia de entre 15º y 45º desde la horizontal del eje, divergente u horizontal, más o menos horizontal con un ángulo de divergencia de entre 0º y 15º hacia arriba o hacia abajo desde la horizontal del eje (también llamado divaricado), reclinado, direccionado hacia abajo, con un ángulo de divergencia de entre 15º y 45º desde la horizontal del eje, descendiente, direccionado hacia abajo, con un ángulo de divergencia de entre 15º y 45º con respecto a la parte baja del eje, y deprimido, apretado cercanamente al eje hacia abajo, con un ángulo de divergencia de entre 0º y 15º con respecto a la parte baja del eje.
Otros términos de orientación son más generales. Antrorso significa doblado hacia o direccionado hacia arriba, usualmente hace referencia a pequeños apéndices, retrorso significa doblado hacia o direccionado hacia abajo. Connivente significa convergente apicalmente sin fusión, como en las anteras que se ponen juntas en la flor (como en Solanum). Erecto es apuntando hacia arriba (usualmente sin hacer referencia a un eje). Pendiente o péndulo significa que cuelga. Deflejo significa doblado abruptamente hacia abajo. Reflejo significa doblado hacia abajo.
Postura y prefoliación
editarLa postura hace referencia a la ubicación en relación con un plano. Puede ser posteriormente clasificada como transversa, longitudinal, doblada. La ptyxis o vernación o prefoliación se refiere a la postura de las estructuras durante su desarrollo, cuando todavía no llegaron a la madurez, como la de las hojas inmaduras o las piezas de un pimpollo. Muchos de los mismos términos usados para postura de órganos maduros pueden ser usados para la vernación. En este texto se considerarán al mismo tiempo.
La postura transversa es la ubicación de la punta (la parte distal) de un objeto con respecto al plano en el que comenzó su desarrollo. Términos de postura transversa son recurvo, con la punta gradualmente curvándose hacia afuera o hacia abajo (abaxialmente), cernuosa, con la punta cayéndose hacia abajo (abaxialmente), escuarrosa, fuertemente curvada cerca del ápice hacia abajo o hacia afuera (abaxialmente), como en los filarios de algunas Asteraceae, incurva, con la punta gradualmente curvándose hacia adentro o hacia arriba (adaxialmente), plana o recta, sin curvas, y flexuosa, con el eje central y la punta doblándose alternativamente hacia arriba y hacia abajo.
-
 Postura recurva.
Postura recurva. -
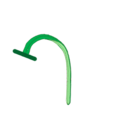 Postura cernuosa.
Postura cernuosa. -
 Postura escuarrosa.
Postura escuarrosa.



-
 Postura incurva.
Postura incurva. -
 Postura flexuosa.
Postura flexuosa.


La postura longitudinal es la ubicación de los márgenes de un objeto con respecto al plano en el que comenzó su desarrollo. Términos comunes son conduplicado, longitudinalmente doblado en un eje central, con los lados adaxiales adyacentes mirándose cara a cara (la prefoliación conduplicada, con el limbo plegado en dos a lo largo de la vena media, como en Petunia thymifolia), revoluto, con el margen o la porción más externa de los lados enrollándose hacia afuera o hacia abajo sobre la superficie abaxial (la prefoliación revoluta, con cada mitad del limbo enrollada hacia el lado abaxial hasta la vena media, como en el sauce y en Muehlenbeckia sagittifolia), involuto, con los márgenes o la porción más externa de los lados enrollándose hacia adentro o hacia arriba hacia el lado adaxial (la prefoliación involuta, con cada mitad del limbo enrollada hacia el lado adaxial hasta la vena media, como en el álamo y el peral), en forma de copa, cóncavo o convexo a lo largo de toda la superficie (abaxialmente o bien adaxialmente), replegado, con una serie de dobleces en el eje longitudinal (la prefoliación replegada, con el limbo plegado como un abanico, como en la vid) (subcategorías son induplicado, replegado con los lados adaxiales adyacentes mirándose unos a otros, con forma de V en el plano transversal, y reduplicado, replegado con los lados abaxiales adyadentes mirándose unos a otros, con forma de V invertida en el plano transversal, la prefoliación reduplicada, doblada a lo largo de la vena media con los lados abaxiales mirándose[5]). Undulado o repandido, con los márgenes en ondas en el plano vertical. La prefoliación convoluta o prefoliación supervoluta, con el limbo enrollado paralelamente a la vena media, una mitad enrollada dentro de la otra, como en la lechuga y miembros de Zingiberales.
-
 Postura longitudinal conduplicada, o prefoliación conduplicada. Corte transversal. 1: Haces vasculares de la vena media. 2: Tallo.
Postura longitudinal conduplicada, o prefoliación conduplicada. Corte transversal. 1: Haces vasculares de la vena media. 2: Tallo. -
 Postura longitudinal revoluta, o prefoliación revoluta. Corte transversal. 1: Haces vasculares de la vena media. 2: Tallo
Postura longitudinal revoluta, o prefoliación revoluta. Corte transversal. 1: Haces vasculares de la vena media. 2: Tallo -
 Postura longitudinal revoluta en hojas de Cycas revoluta.
Postura longitudinal revoluta en hojas de Cycas revoluta. -
 Postura longitudinal involuta, o prefoliación involuta. Corte transversal. 1: Haces vasculares de la vena media. 2: Tallo
Postura longitudinal involuta, o prefoliación involuta. Corte transversal. 1: Haces vasculares de la vena media. 2: Tallo




-
 Prefoliación involuta en el loto (Nelumbo).
Prefoliación involuta en el loto (Nelumbo). -
 Prefoliación involuta en pétalos de Cucurbita. Flor durante su apertura.
Prefoliación involuta en pétalos de Cucurbita. Flor durante su apertura. -
 Postura longitudinal con forma de copa. Corte transversal. 1: Haces vasculares de la vena media. 2: Tallo.
Postura longitudinal con forma de copa. Corte transversal. 1: Haces vasculares de la vena media. 2: Tallo. -
 Postura longitudinal replegada.
Postura longitudinal replegada.

_flor_masculina_M01_antesis_vista_superior_p%C3%A9talos_desenroll%C3%A1ndose_.JPG)


-
 Prefoliación replegada. Corte transversal. 1: Haces vasculares de la vena media. 2: Haces vasculares de las venas secundarias.
Prefoliación replegada. Corte transversal. 1: Haces vasculares de la vena media. 2: Haces vasculares de las venas secundarias. -
 Prefoliación reduplicada. Corte transversal. 1: Haces vasculares de la vena media. 2: Tallo.
Prefoliación reduplicada. Corte transversal. 1: Haces vasculares de la vena media. 2: Tallo. -
 Prefoliación convoluta o supervoluta. Corte transversal. 1: Haces vasculares de la vena media. 2: Tallo
Prefoliación convoluta o supervoluta. Corte transversal. 1: Haces vasculares de la vena media. 2: Tallo



Otros tipos de vernación incluyen: Plana, con el limbo extendido. Doblada, con el limbo plegado en dos a lo largo de una línea transversal.[69] Circinada, con la lámina (incluyendo el raquis y la raquilla si presente) enrollado del ápice a la base, como en las hojas de muchos helechos y cícadas.
-
 Prefoliación doblada. 1: Haces vasculares de la vena media.
Prefoliación doblada. 1: Haces vasculares de la vena media. -
 Prefoliación circinada. 1: Haces vasculares de la vena media.
Prefoliación circinada. 1: Haces vasculares de la vena media.


La postura de twisting/bending se refiere a la postura de un objeto que se está torciendo en relación al plano en el que comienza su desarrollo. Resupinado significa girado 180º, como en las hojas de Alstroemeriaceae o los ovarios de la mayoría de las flores de Orchidaceae. Geniculado significa con una postura en zig-zag, como en el raquis de la inflorescencia de algunos Poaceae. Voluble (twining) es enroscado alrededor de un eje central, como en muchas enredaderas. Los tallos de las enredaderas pueden estar girados de forma dextrorsa, doblándose helicoidalmente como un típico tornillo de mano derecha, como en algunas Convolvulaceae, o sinistrorsa, como un tornillo de mano izquierda, como en algunas Caprifoliaceae.
Superficie
editarNumerosos términos describen la superficie de órganos o piezas de la flor. Los caracteres de la superficie pueden dividirse en tres tipos de caracteres: la configuración, la excrecencia de la epidermis, y la vestitura o indumento. Además pueden ser descriptos el tipo de tricoma y de cerdas como caracteres de superficie.
Configuración
editarLa configuración se refiere a los patrones gruesos de las células de la epidermis que no sean causados por la venación o las excrecencias. Estos incluyen canaliculado, con surcos longitudinales, usualmente en relación con pecíolos o venas medias, fenestrado, con hoyos como ventanas en la superficie como en Monstera deliciosa, Araceae, punteado, cubierto con depresiones diminutas como hoyitos, rugoso, cubierto por líneas reticuladas burdas, usualmente con áreas como ampollas entre ellas, ruminado, de textura desigual, bastamente arrugado, que se ve como masticado, suave o plano, con una configuración suave, estriado, con líneas finas longitudinales, arrugado (wrinkled), con líneas finas o deformaciones finas, irregulares.
Excrecencia de la epidermis
editarLa excrecencia de la epidermis se refiere a los patrones de superficie causados por secreciones o crecimientos estructurales de la epidermis que no sean tricomas o cerdas (que se verán a continuación). Términos son:
- glandular, cubierto con glándulas diminutas negruzcas o translúcidas (no tricomas glandulares),
- glauco, cubierto con una capa de ceras suave y usualmente blancuzca (que puede ser quitada al tocarse),
- nítido, con apariencia de lustroso, y glutinoso o víscido, con una superficie brillante y pegajosa.
Términos que denotan crecimientos de la epidermis son
- acúleo o aguijoneado, con crecimientos espinosos llamados aguijones (no son espinas, ver en aguijón (botánica)),
- farináceo, cubierto con pequeños gránulos,
- papiloso, tuberculado o verrucado, con diminutas proturberancias redondeadas,
- escabroso, con una superficie rugosa, como la del papel de lija.
Vestitura/indumento
editarVestitura, llamada a menudo indumento, denota la cubierta de tricomas. Los tricomas son estructuras como pelos de la superficie que pueden funcionar para proteger a la planta de la herbivoría, en reflejar la luz visible y la ultravioleta, y en inhibir la transpiración de agua. Los términos de la vestitura son una combinación de tipo de tricoma, largo, fortaleza, forma, densidad y color.
Glabro es el término que significa sin tricomas en absoluto. Subglabro significa casi glabro (con sólo unos pocos tricomas esparcidos), glabrado (glabrate) puede significar lo mismo o puede decir que se va volviendo glabro con el tiempo por la pérdida de tricomas.
Barbado (bearded) significa con un único parche de tricomas que se eleva desde la superficie de un objeto, por ejemplo desde un pétalo. Comoso es similar, pero se refiere a un parche apical de tricomas, por ejemplo en una semilla. Dos términos que también son tratados en "margen" son ciliado, con tricomas marginales conspicuos, y ciliolado, con pequeños tricomas marginales.
Pubescente es un término común que significa con tricomas más o menos derechos, cortos, suaves, un poco esparcidos, y delgados. También puede ser usado como un término general, significando "que tiene tricomas". Puberulento significa diminutamente pubescente, es decir pubescente con tricomas muy cortos o muy esparcidos. Canescente o incano significa pubescente y blancuzco, cubierno con tricomas densos y finos grisáceos a blancos. Tomentoso significa cubierto por tricomas muy densos imbricados. Tomentuloso significa diminutamente tomentoso, es decir tomentoso con los tricomas muy cortos.
Villoso o lanado significa cubierto con tricomas largos, suaves y sinuosos, si bien lanado tiene tricomas más cortos que villoso, estos términos se intergradan y probablemente sea mejor tratarlos como sinónimos. Piloso significa con tricomas suaves, rectos a ligeramente enmarañados, generalmente a ángulos rectos de la superficie. Aracnoso o aracnoide significa con tricomas formando una masa densa, como una telaraña (pero parece villoso y puede ser confundido con ese tipo).
Escabroso significa con tricomas ásperos, como los del papel de lija, escaberuloso significa diminutamente escabroso (escabroso también es un tipo de excrecencia epidérmica porque la escabrosidad puede ser debida tanto a tricomas como a crecimientos de la epidermis). Hirsuto significa con tricomas largos, bastante rígidos (pero que no penetran la piel), híspido significa con tricomas bastante largos y rígidos que muchas veces son capaces de penetrar la piel. El término urento significa con tricomas híspidos que son urticantes, como en Urtica, la ortiga.
Estrigoso es cubierto por tricomas densos, toscos, curvados, y mayormente aplanados, muchas veces con una base bulbosa. Estriguloso significa diminutamente estrigoso, es decir con la misma morfología pero un tamaño mucho menor. Seríceo significa con tricomas largos y apretados que tienen una apariencia de seda o lustrosa.
Flocoso significa con tricomas densos en muchos parches. Lepidoto significa cubierto con escamas o estructuras como escamas, es un carácter que se intergrada con los de excrecencias epidérmicas.
Tipos de tricomas y cerdas
editarLos tricomas o pelos son pequeños apéndices de la epidermis. El tipo de tricoma se refiere a la estructura específica microscópica de tricomas individuales y puede estar dentro del campo de la anatomía de plantas. Constituye un valioso carácter sistemático. Si bien el tipo de tricoma puede estar correlacionado con la vestitura, vestitura hace referencia a la apariencia al ojo desnudo de las masas de tricoma. Por ejemplo una vestitura tomentosa puede tener un número de tipos de tricoma, por ejemplo dendríticos, estrellados, etc.
El tipo de tricoma puede tener en cuenta el número de células que forman el tricoma. Un tricoma unicelular es el que consiste en una única célula y usualmente es muy pequeño. Un tricoma multicelular contiene dos o más células. Los tricomas multicelulares pueden ser uniseriados, con una única línea vertical de células, o multiseriados, con más de una línea vertical de células. El número de capas de células en un tricoma también puede ser un carácter diagnóstico.
-
 Tricoma unicelular.
Tricoma unicelular. -
 Otros tipos de tricomas unicelulares.
Otros tipos de tricomas unicelulares. -
 Tricoma multicelular uniseriado (en este caso con punta aguzada).
Tricoma multicelular uniseriado (en este caso con punta aguzada).



-
 Tricoma multicelular biseriado.
Tricoma multicelular biseriado. -
 Tricoma multicelular multiseriado (en este caso con punta aguzada).
Tricoma multicelular multiseriado (en este caso con punta aguzada).


Muchos tricomas son diagnosticados según su forma general y morfología. Los tricomas aguzados ("tapering") son los que terminan en una punta aguzada. Los tricomas malpighianos o con forma de T son los que tienen dos brazos que se elevan desde una base común, como en muchos miembros de Malpighiaceae. También hay tricomas con forma de Y.[5] Los tricomas glandulares son tricomas secretorios o excretorios, usualmente con una célula glandular apical. Los tricomas glandulares pueden ser pilados-glandulares, con una célula glandular en la punta de un eje basal, o capitados-glandulares, con una célula glandular con un eje basal muy corto o sin eje basal. Estos son sólo dos esquematizaciones, hay muchas variaciones de los tricomas glandulares, que se observan en las imágenes de abajo.[5] Los tricomas ramificados pueden ser de dos tipos: estrellados, que son tricomas con forma de estrella con muchos brazos que se elevan de una base común (que puede tener eje o ser sésil), y dendríticos, que son tricomas como árboles con múltiples ramas laterales. De nuevo hay muchos tipos de tricomas ramificados: los hay con ramas largas, con ramas cortas.[5] Hay tricomas en forma de pelos agrupados.[5] Los tricomas peltados son los que tienen una porción apical como un disco en la punta de un eje. Las escamas sésiles[5] son tricomas multiseriados con una lámina de una célula de espesor.
-
Tricoma aguzado uniseriado.
-
 Tricomas uniseriados en Coleus, foto al microscopio (SEM). Se observa uno glandular, ver más adelante.
Tricomas uniseriados en Coleus, foto al microscopio (SEM). Se observa uno glandular, ver más adelante. -
Tricoma aguzado multiseriado.
-
 Tricoma malpighiano o con forma de T.
Tricoma malpighiano o con forma de T.


-
 Tricoma con forma de Y.
Tricoma con forma de Y. -
 Tricoma pilado-glandular. En verde oscuro la glándula.
Tricoma pilado-glandular. En verde oscuro la glándula. -
 Tricoma capitado-glandular. En verde oscuro la glándula.
Tricoma capitado-glandular. En verde oscuro la glándula. -
 Tricoma glandular con cabeza de 1 célula. En verde oscuro la glándula.
Tricoma glandular con cabeza de 1 célula. En verde oscuro la glándula.




-
 Tricoma glandular con cabeza de 2 células, en este caso casi sésil. En verde oscuro la glándula.
Tricoma glandular con cabeza de 2 células, en este caso casi sésil. En verde oscuro la glándula. -
 Tricoma glandular con cabeza multicelular. En verde oscuro la glándula.
Tricoma glandular con cabeza multicelular. En verde oscuro la glándula. -
 Tricoma con cabeza glandular multicelular con eje biseriado. En verde oscuro la glándula.
Tricoma con cabeza glandular multicelular con eje biseriado. En verde oscuro la glándula. -
 Tricoma con cabeza glandular multicelular y eje multiseriado. En verde oscuro la glándula.
Tricoma con cabeza glandular multicelular y eje multiseriado. En verde oscuro la glándula.




-
 Tricoma glandular de Juglans nigra, foto al microscopio (SEM).
Tricoma glandular de Juglans nigra, foto al microscopio (SEM). -
 Tricomas glandulares en flor de Nicotiana alata, foto SEM.
Tricomas glandulares en flor de Nicotiana alata, foto SEM. -
 Tricoma glandular pero con la glándula en célula basal modificada. En verde oscuro la glándula.
Tricoma glandular pero con la glándula en célula basal modificada. En verde oscuro la glándula. -
 Tricoma glandular unicelular, con glándula en la base (rodeada de una base de células epidérmicas más pequeñas) en la ortiga, Urtica dioica. En este caso el tricoma se rompe, liberando los compuestos tóxicos producidos por la glándula, al ser tocado.
Tricoma glandular unicelular, con glándula en la base (rodeada de una base de células epidérmicas más pequeñas) en la ortiga, Urtica dioica. En este caso el tricoma se rompe, liberando los compuestos tóxicos producidos por la glándula, al ser tocado.




-
 Tricomas de Urtica dioica.
Tricomas de Urtica dioica. -
 Tricoma estrellado sésil.
Tricoma estrellado sésil. -
 Tricoma estrellado con eje.
Tricoma estrellado con eje. -
 Tricoma estrellado con eje en Arabidopsis thaliana, foto al microscopio (SEM).
Tricoma estrellado con eje en Arabidopsis thaliana, foto al microscopio (SEM).




-
 Tricoma dendrítico.
Tricoma dendrítico. -
 Tricoma dendrítico, otro tipo.
Tricoma dendrítico, otro tipo. -
 Tricoma con ramas cortas.
Tricoma con ramas cortas. -
 Tricomas dendríticos en la hoja de lavanda, Lavandula.
Tricomas dendríticos en la hoja de lavanda, Lavandula.




-
 Tricoma en forma de pelos agrupados.
Tricoma en forma de pelos agrupados. -
 Tricomas dendríticos en una planta "pitcher". Foto al microscopio (SEM).
Tricomas dendríticos en una planta "pitcher". Foto al microscopio (SEM). -
 Tricoma peltado.
Tricoma peltado. -
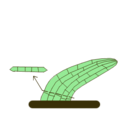 Escama sésil.
Escama sésil.




Los tricomas también pueden ser delimitados según su posición y función. Por ejemplo, los tricomas encontrados en las axilas de hojas típicamente envainadoras, que pueden funcionar en secretar mucílago protector, se llaman escuámulos intravaginales (o axilares), como se encuentra en muchos Alismatales.
Las cerdas ("bristles") son similares a los tricomas pero son en general mucho más robustas, si bien las cerdas y los tricomas pueden intergradarse. Algunas llamadas cerdas son en realidad hojas modificadas, como los gloquidios de Cactaceae. Algunos tipos principales de cerda son barbada o barbelada, con apéndices ("barbas") diminutas, laterales y agudas que pueden tener orientación antrorsa o retrorsa, que se elevan a lo largo de toda la superficie de la cerda, plumosa, como una pluma, cubierta por apéndices ciliados finos y elongados, uncinada o en gancho ("hooked") con una estructura apical como un gancho, y gloquidiada, con estructuras como barbas agrupadas en la punta apical.
-
 Diferenciación entre cerdas (las pequeñas y robustas) y tricomas (como pelos) en la foto al microscopio de una espiguilla de Hordeum murinum. Estas cerdas son simples, sin adornos.
Diferenciación entre cerdas (las pequeñas y robustas) y tricomas (como pelos) en la foto al microscopio de una espiguilla de Hordeum murinum. Estas cerdas son simples, sin adornos.

-
 Una cerda barbada o barbelada.
Una cerda barbada o barbelada. -
 Una cerda plumosa.
Una cerda plumosa. -
 Una cerda uncinada o en gancho.
Una cerda uncinada o en gancho. -
 Cerda gloquidiada (también e incorrectamente, puede verse como cerda gloquidiada al gloquidio de Cactaceae, que es una hoja modificada y por lo tanto es una espina foliar con barbas retrorsas en su ápice).
Cerda gloquidiada (también e incorrectamente, puede verse como cerda gloquidiada al gloquidio de Cactaceae, que es una hoja modificada y por lo tanto es una espina foliar con barbas retrorsas en su ápice).




Simetría
editarSi bien el término usualmente es utilizado en referencia a las flores, puede ser un carácter general para describir cualquier órgano o pieza. La simetría es definida por la presencia y número de imágenes especulares o planos de simetría. La simetría zigomórfica o bilateral (también llamada irregular) es la que tiene un único plano de simetría, dividiendo al órgano en dos imágenes especulares. La simetría birradial es la que tiene dos y sólo dos planos de simetría. La simetría actinomórfica o radial (también llamada regular) es la que tiene tres o más planos de simetría, que se unen en el centro. Asimétrico significa sin planos de simetría. A veces la distinción entre radial y birradial no es hecha, llamándose a las dos simetría radial o actinomorfía, sin embargo la distinción es útil y por eso es reconocida aquí.
Fenómenos temporales
editarLos fenómenos temporales tienen en cuenta cualquier consideración específica del tiempo. Se dividen en duración, maduración, y perioricidad.
La duración es el tiempo de vida de una planta o parte de una planta. La duración de una planta describe el largo de la vida de una planta, las categorías más comunes son anual, bienal, perenne. Fueron descriptas en la sección #Hábito, duración, hábitat y distribución
Otros términos de duración describen a las partes de la planta, por ejemplo para las hojas. Perenne o siempreverde ("evergreen") significa persistente por dos o más estaciones de crecimiento, como en las hojas de la mayoría de las coníferas. Deciduo significa con las partes persistentes por una estación de crecimiento y luego cayendo, como en las hojas de Acer. Deciduo también se aplica a las partes de la flor que se caen después de la fecundación.[70] Efímero o fugaz significa que se cae muy temprano (en comparación a lo que es típico) y usualmente se aplica a las partes florales. Persistente se aplica a las partes de la flor que quedan acompañando al fruto, como el cáliz de Physalis y de Malus.[70] Marcescente significa efímero pero con restos persistentes, es decir los restos se marchitan pero son persistentes, como las corolas que quedan adheridas durante la formación del fruto. También son marcescentes las hojas que se marchitan pero no caen durante la estación desfavorable.[5] Acrescente se refiere a las partes de la planta que persisten y continúan creciendo más allá de lo típico, como el cáliz de Physalis (Solanaceae), que se expande considerablemente y funciona como una parte accesoria que encierra el fruto. Monocárpico se dio en la sección #Hábito, duración, hábitat y distribución.
-
 El aguacate o palta, Persea americana, posee un fruto con tépalos persistentes: se mantienen pero no se desarrollan durante la formación del fruto.
El aguacate o palta, Persea americana, posee un fruto con tépalos persistentes: se mantienen pero no se desarrollan durante la formación del fruto. -
 La castaña es el fruto de Castanea sativa (una fagácea), con pistilo y estilos persistentes.
La castaña es el fruto de Castanea sativa (una fagácea), con pistilo y estilos persistentes. -
 Los robles (Quercus) poseen hojas marcescentes: se marchitan pero pasan todo el invierno unidas al árbol, completando la abscisión recién en la primavera siguiente.
Los robles (Quercus) poseen hojas marcescentes: se marchitan pero pasan todo el invierno unidas al árbol, completando la abscisión recién en la primavera siguiente. -
 El cáliz de la solanácea (tomates y papas) Physalis es acrescente: se sigue desarrollando durante la formación del fruto, en este caso envolviendo a la parte comestible en una cubierta papirácea.
El cáliz de la solanácea (tomates y papas) Physalis es acrescente: se sigue desarrollando durante la formación del fruto, en este caso envolviendo a la parte comestible en una cubierta papirácea.


_during_Autumn_along_Terrace_Boulevard_in_Ewing,_New_Jersey.jpg)

Maduración hace referencia al tiempo relativo del desarrollo de las partes de la planta. El término antesis hace referencia al momento de la floración, cuando las flores se abren con sus partes disponibles para la polinización. Protándrico (que significa "el macho primero") quiere decir que los estambres o anteras se desarrollan antes que los carpelos o el estigma. Protoginio ("la hembra primero") significa que el estigma o los carpelos maduran antes que los estambres o las anteras. Los dos son mecanismos para promover la fecundación cruzada dentro de una especie.
La perioricidad hace referencia a los fenómenos que se repiten periódicamente. Términos que hacen referencia al momento del día son diurnos (durante el día), nocturnos (durante la noche), matutinos (durante la mañana), y vespertinos (durante el atardecer). Estos términos son utilizados usualmente en referencia a cuándo se abren las flores de un taxón dado. Otros términos se corresponden a las estaciones, como vernal (que aparece en la primavera), estival (que aparece en el verano), u otoñal (que aparece en el otoño).
Glosario de Cactaceae
editarUn tipo muy especializado de vástago deterrente de herbívoros son las areolas, formadas por un vástago apical modificado, reducido, que no se elonga y de donde se originan las espinas foliares (definidas como estructuras agudas con tejido vascular en su interior, provenientes de una hoja o parte con tejido vascular de una hoja[71]). Las areolas son características de la familia de los cactus (Cactaceae).
- Cactaceae
-
 Areola de Cactaceae. En amarillo claro el vástago, en verde espinas foliares, en marrón un segundo tipo de espinas foliares encontrado en cactus opuntioides llamado gloquidio (véase en espina foliar).
Areola de Cactaceae. En amarillo claro el vástago, en verde espinas foliares, en marrón un segundo tipo de espinas foliares encontrado en cactus opuntioides llamado gloquidio (véase en espina foliar). -
 Detalle del cladodio de Opuntia basilaris mostrando las areolas con gloquidios y espinas foliares.
Detalle del cladodio de Opuntia basilaris mostrando las areolas con gloquidios y espinas foliares.


Glosario de Poaceae
editar.svg)
.svg)

El culmo se refiere al sector de los tallos que no forman parte del rizoma subterráneo, sino que emergen de la tierra y da flores y frutos en los pastos de poáceas. El término también se usa para la familia de plantas graminiformes Cyperaceae. Los culmos leñosos se llaman cañas, como las de las bambusóideas.
Un macollo (tiller) es un término general para un vástago proliferativo (que finaliza en un culmo) de las gramíneas, que nace en la axila de la hoja en la base de otro culmo o del tallo rizomatoso, y debido a que no tiene crecimiento secundario, en su primer entrenudo es del diámetro de la yema, a medida que se adicionan entrenudos aumentan de diámetro y además se originan en estos nudos raíces adventicias, hasta que a cierta distancia del culmo anterior, que depende de la especie, el tallo se vuelve erguido (ortótropo) y emerge de la tierra como un culmo nuevo, dejando un "cuello" en su inserción con el tallo anterior. Al culmo en cuya base se origina otro culmo se llama "culmo macollante" (tillering culm). Son macollos intravaginales los que permanecen cerca de su origen, y extravaginales los que se elongan, los primeros generan pastos cespitosos y los segundos pastos de hábito "abierto" apto para cobertura del suelo. ([4]:182)
Una espiguilla de Poaceae consiste en un eje (raquilla) típicamente con dos brácteas basales (glumas) y una o más unidades que son ramas cortas laterales (el antecio), cada una con dos brácteas (la lemma y la palea), que tienen una flor terminal única reducida.
Las brácteas especializadas de la espiguilla de Poaceae tienen diferentes nombres: gluma y glumela son las dos brácteas que se encuentran en la base de la espiguilla (inflorescencia), la lemma y la pálea son las dos brácteas que se encuentran por debajo de cada flor individual de la espiguilla.
- Espiguilla de Poaceae
-
 En Poaceae hay 4 términos para brácteas específicas, la gluma y la glumela están por debajo de toda la inflorescencia, mientras que la lemma y la pálea están por debajo de cada flor individual.
En Poaceae hay 4 términos para brácteas específicas, la gluma y la glumela están por debajo de toda la inflorescencia, mientras que la lemma y la pálea están por debajo de cada flor individual. -
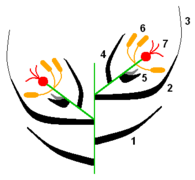 Espiguilla de Poaceae, esquema.
Espiguilla de Poaceae, esquema.


El perianto es altamente modificado para una función no relacionada con protección o atracción, reducido a dos lodículas que al modificarse abren las brácteas y exponen los estambres o estigmas de la flor.
Glosario de Asteraceae
editar



_protecting_the_head.JPG)
Lo que parece una flor en margaritas y girasoles (:Asteraceae, =Compositae) es en realidad una inflorescencia (antiguamente llamada "flor compuesta", compuesta por diminutas flores, sinónimo del término todavía utilizado pseudanto), las diminutas flores por convención clasificadas en discos y rayos. La flor llamada rayo, en general ubicada en los márgenes de la cabeza, tiene perianto ligulado o también llamado rayo: una corola corta y tubular con una única extensión apical acintada (el rayo), cada una de estas flores tiene el aspecto de un "pétalo". La flor llamada disco en referencia a que suele ser encontrado dentro del disco (en el interior) de la inflorescencia, tiene perianto discoide que consta de una corola tubular gamopétala con 5 lóbulos acampanados que representan el ápice de sus 5 pétalos.
- Compuestas
-
 Capítulo: flores con perianto discoide (interiores), rayos (externas). Del interior al exterior y de arriba abajo: Blanco, ápice del gineceo (el estilo con dos ramas). En amarillo fuerte: ápice de los estambres (anteras unidas formando un tubo). Amarillo claro y naranja: perianto gamopétalo (incluyendo la larga lígula del rayo). Verde oscuro: filarios.
Capítulo: flores con perianto discoide (interiores), rayos (externas). Del interior al exterior y de arriba abajo: Blanco, ápice del gineceo (el estilo con dos ramas). En amarillo fuerte: ápice de los estambres (anteras unidas formando un tubo). Amarillo claro y naranja: perianto gamopétalo (incluyendo la larga lígula del rayo). Verde oscuro: filarios. -
 Corte longitudinal. Verde agua: receptáculo. Verde oscuro: involucro de filarios. Se observa el ovario ínfero justo debajo de la corola gamopétala.
Corte longitudinal. Verde agua: receptáculo. Verde oscuro: involucro de filarios. Se observa el ovario ínfero justo debajo de la corola gamopétala. -
 Estambres unidos por sus anteras formando un tubo, estambres sinantéreos (= de anteras singenesias) internos.
Estambres unidos por sus anteras formando un tubo, estambres sinantéreos (= de anteras singenesias) internos.


Estas pequeñas flores a veces tienen por debajo brácteas que han recibido diferentes nombres, como pálea (sensu Linneo; también llamadas "escamas axilantes", "escamas receptaculares", "brácteas interseminales",... en autores posteriores[72] y en algunos glosarios "flósculo" (nombre que también comparte a veces con las espiguillas de las gramíneas).
Otras asteráceas tienen solo flores liguladas (rayos), como Taraxacum (diente de león) en el que el papus o vilano (cáliz), en forma de pelos acrescentes, crece junto con el fruto y forma parte de él.
Las brácteas debajo de las inflorescencias son las brácteas involucrales o filarios, que en conjunto forman el involucro, con función sepaloide.
El estilo puede ser separado del estigma por un anillo colector que actúa como cepillo para arrastrar el polen en su paso por el tubo estaminal, por ejemplo en la tribu Cardueae de la familia Asteraceae, donde el estilo está encerrado en el tubo formado por las anteras soldadas de los 5 estambres. De esta manera, el polen se concentra en la base de las 2 ramas estigmáticas para facilitar la autofecondación.
-
 Flores de perianto discoide en una Leucanthemum vulgare
Flores de perianto discoide en una Leucanthemum vulgare -
 Detalle del perianto discoide, los estambres sinantéreos y el estilo con sus dos ramas en L. vulgare.
Detalle del perianto discoide, los estambres sinantéreos y el estilo con sus dos ramas en L. vulgare. -
 Nombres de las partes en un Helianthus bolanderi.
Nombres de las partes en un Helianthus bolanderi.



Glosario de Fabaceae
editar
Las leguminosas tienen una flor y fruto característicos con terminología específica de ese taxón. El fruto es una "legumbre" en forma ancestral y está bien conservada en el ovario y el fruto inmaduro, al madurar diverge en diferentes tipos de fruto con diferentes tipos de dispersión.
El perianto de la subfamilia Faboideae es papilionáceo, con un pétalo posterior grande, dos internos y laterales, y dos pétalos inferiores usualmente apicalmente conados.
La prefloración es vexilar. Hay un pétalo externo y superior (el estandarte o vexilo), que abraza los laterales llamados alas, y éstas cubren a su vez a los inferiores soldados entre sí, que forman la quilla. Característica de la subfamilia Papilionoideae.
En el ovario de la flor se pueden observar los caracteres que indican que al fructificar será una legumbre o uno de sus derivados. Una legumbre (o vaina, a veces "vaina" se usa de una forma más general para indicar morfología y entonces también es vaina el fruto de la vainilla, que es una orquídea) es un fruto seco y dehiscente derivado de un único carpelo, que se abre a lo largo de dos suturas longitudinales ("dehiscencia sutural doble"), las legumbres son el fruto ancestralmente de todas las leguminosas, Fabaceae o Leguminosae. Algunas legumbres retienen el vestigio de dos suturas longitudinales, pero han sido secundariamente modificadas, como los lomentos, que se abren transversalmente en segmentos de una sola semilla (ejemplos son Aeschynomene, Desmodium, Sophora, Hedysarum[5]), y las legumbres indehiscentes, que no se abren en absoluto (como el maní). Algunos autores diferencian la legumbre indehiscente de maduración aérea de la legumbre indehiscente de maduración subterránea, a la que llaman geocarpo.
-
 Legumbre, fruto cerrado, fruto abierto y corte transversal.
Legumbre, fruto cerrado, fruto abierto y corte transversal. -
 Lomentos madurando en Hedysarum.
Lomentos madurando en Hedysarum. -
 Lomentos madurando en Hedysarum, se observan las líneas de dehiscencia.
Lomentos madurando en Hedysarum, se observan las líneas de dehiscencia. -
 Legumbre indehiscente: Arachis hypogaea.
Legumbre indehiscente: Arachis hypogaea.


.jpg)

En las semillas puede ser importante, en particular para la identificación de los géneros dentro de la familia, la presencia o ausencia en el tegumento de una fina marca en forma de anillo o de herradura en medio de cada lado, la línea fisural o pleurograma[73][74] que rodea la areola lateral de la semilla.
Glosario de Cucurbitaceae
editarLas cucurbitáceas tienen una flor y fruto característicos, y el ovario de la flor, y el fruto que madura a partir de él, es un pepónide.
Un pepónide (pepo) es un fruto carnoso, ancestralmente es el de todas las cucurbitáceas (melón, sandía, pepino, calabazas, etc.), es macizo (no hueco) desde su etapa de ovario, chequear: sin septos con placentación parietal (las placentas intrusas normalmente ocupando todo el interior del ovario y del fruto inmaduro y luego maduro), de exocarpio coriáceo y mesocarpio carnoso, el endocarpio representado por solo una membranita que rodea a las semillas.
El pepónide es derivado de un ovario ínfero, por lo que deben observarse en el extremo distal las cicatrices que dejaron el perianto y el androceo al caer.
Las semillas son aplanadas y dispuestas en un plano transversal al eje del fruto.
El pepónide se determina con más facilidad en los estadios de ovario y las primeras etapas de maduración del fruto, donde se observa su ovario macizo (no hueco) y la disposición particular de las placentas y las semillas.
- Pepónide
-
 Cucurbita. Ovario. Corte transversal.
Cucurbita. Ovario. Corte transversal. -
 Fruto inmaduro ("zapallito").
Fruto inmaduro ("zapallito"). -
 Fruto cosechado muy tempranamente maduro.
Fruto cosechado muy tempranamente maduro.
_ovario,_corte_transversal.jpg)

_2015-01-29_zapallo_d%C3%ADa_19,_pulpa_y_semillas.jpg)
-
 Cucurbita. Ovario.
Cucurbita. Ovario. -
 Pepónide y corte transversal. Exocarpio coriáceo, mesocarpio carnoso, y cicatriz en el ápice que dejaron el perianto y el androceo.
Pepónide y corte transversal. Exocarpio coriáceo, mesocarpio carnoso, y cicatriz en el ápice que dejaron el perianto y el androceo. -
 Corte transversal del fruto de Luffa, exocarpio removido, se mantiene solo el mesocarpio fibroso.
Corte transversal del fruto de Luffa, exocarpio removido, se mantiene solo el mesocarpio fibroso. -
 Corte de Luffa.
Corte de Luffa.


.JPG)
Glosario de Arecaceae
editarTres términos específicos de inflorescencias de palmeras son: infrafoliar, en que la inflorescencia aparece en una yema axilar debajo de la corona de hojas, interfoliar, cuando aparece en medio de la corona de hojas, y suprafoliar, cuando aparece por arriba de la corona de hojas.
Glosario de Brassicaceae
editarLas silicuas y las silículas son frutos secos y dehiscentes derivados de un ovario de dos carpelos que se abre a lo largo de dos suturas pero tiene una partición persistente, el "replum" (el septo maduro, en general con las semillas adheridas, "dehiscencia placentífraga"). Los dos tipos de fruto difieren en que la silícula es tan ancha o más ancha que larga, y la silicua es más larga que ancha. Los dos son los tipos característicos de frutos de Brassicaceae.
-
 Silicua, fruto cerrado y fruto abierto y corte transversal.
Silicua, fruto cerrado y fruto abierto y corte transversal. -
 Silicuas de Cardamine impatiens.
Silicuas de Cardamine impatiens. -
 Silicuas de Rorippa microphylla al momento de la dehiscencia.
Silicuas de Rorippa microphylla al momento de la dehiscencia.


.jpg)
-
 Silícula.
Silícula. -
 Silículas de Lunaria annua. Se observa el replum con las semillas adheridas.
Silículas de Lunaria annua. Se observa el replum con las semillas adheridas.


Glosario de Apiaceae
editarUn esquizocarpo de mericarpios es uno en que los carpelos de un ovario se dividen durante la maduración del fruto, cada carpelo desarrollándose en una unidad mericarpio, como en Apiaceae. Los mericarpios son porciones del fruto que se separan del ovario como una unidad distinta encerrando completamente las semillas, en Apiaceae los dos mericarpios están típicamente unidos entre sí por una estructura como un pequeño eje llamada carpóforo.

.jpg)
Véase también
editarOtros artículos de morfología que complementan a este:
Notas
editar- ↑ Endress (2011[10]): "We can recognize a family from a quick glance at a flower in many cases."
- ↑ a b Ginostemio, ginostegio, polinio, polinario, aparato traslador
(en Orchidaceae y Apocynaceae)
En algunos taxones el androceo y el gineceo están fusionados en una estructura común llamada ginostemio (Orchidaceae) o ginostegio (Apocynaceae), ambos formadores de polinarios (el contenido de todo el polen empaquetado en un polinio más un aparato traslador).
El polinio es una antera típicamente diteca en que todo el polen de las dos tecas (Orchidaceae) o de tecas adyacentes (Asclepias) se fusiona en una única masa. Los polinios de Orchidaceae y Asclepias tienen diferente origen evolutivo y de desarrollo.
- En Asclepias además las anteras se adhieren al estigma formando el ginostegio; el polen de cada teca forma un polinio. En este género, los polinios de dos tecas vecinas se unen entre sí mediante las caudículas y el retináculo que son producciones del estigma. El conjunto de retináculo, caudículas y polinios constituye el aparato translador.[57]
- En Orchidaceae (orquídeas) parece que los estambres y el estilo formasen por concrescencia (por unión congénita) una columna llamada ginostemo, sobre la cual se asienta la única antera, protegida por una cubierta que se desprende, y el estigma.[57]



{kind=link}
Referencias
editar- ↑ a b c d e f g h i j k l m n ñ o p q r s t u v w x y z aa ab ac ad ae af ag ah ai aj ak al Simpson, MG. 2005. Plant Systematics. Elsevier Academic Press.
Simpson, MG. 2010. Plant Systematics, Second Edition. Elsevier Academic Press. - ↑ a b Simpson, MG. 2005. Capítulo 10. Plant Anatomy and Physiology. En: Plant Systematics. Elsevier Academic Press.
- ↑ Radford, A. E., W. C. Dickinson, J. R. Massey, y C. R. Bell. 1974. Vascular Plant Systematics. Harper & Row, Nueva York.
- ↑ a b c d e f g h i j Bell, A. D. 1991. Plant Form, an Illustrated Guide to Flowering Plant Morphology. Oxford University Press, Oxford. Disponible en https://archive.org/details/plantform00adri La segunda edición quizás sea verdadera:
Bell, A. D.; Bryan, A. 2008. Plant Form, an Illustrated Guide to Flowering Plant Morphology. Timber Press. https://books.google.com.ar/books/about/Plant_Form.html?id=SM3khPHXhKEC - ↑ a b c d e f g h i j k l m n ñ o p q r s t u v w x y z aa ab ac ad ae af ag ah ai aj ak al am an añ ao ap aq ar Judd W.S., Campbell C.S., Kellogg E.A., Stevens P.F. & Donoghue M.J., Plant Systematics: A Phylogenetic Approach, Ed. 3rd, Sinauer Associates,Sunderland, MA, 2007 Error en la cita: Etiqueta
<ref>no válida; el nombre «Judd et al. 2007» está definido varias veces con contenidos diferentes - ↑ Judd et al. (2007) p.54, en su glosario de morfología de plantas: "Muchos, si no todos, los términos esquematizados aquí deberían ser considerados meramente puntos convenientes a lo largo de un continuum de variación en la forma. Si bien son útiles para la comunicación, condiciones intermedias serán encontradas". En inglés en el original: "Many, if not all, of the terms outlined here should be considered merely convenient points along a continuum of variation in form. Although they are useful in communication, intermediate conditions will be encountered".
Bajo el nombre "Caracteres tipológicos". Por ejemplo en Rato et al. (2016:Rato, C., Harris, D. J., Carranza, S., Machado, L., & Perera, A. (2016). The taxonomy of the Tarentola mauritanica species complex (Gekkota: Phyllodactylidae): Bayesian species delimitation supports six candidate species. Molecular phylogenetics and evolution, 94, 271-278.): "...la especies han sido identificadas y descriptas utilizando caracteres morfológicos, tanto tipológicos como cuantitativos..." En inglés en el original: "...species have been identified and described using morphological traits, either typological or quantitative..." - ↑ Hawthorne y Lawrence. 2012. Plant Identification: Creating User-Friendly Field Guides for Biodiversity Management. Capítulo 5: «Identification: Keys and other access methods.»
- ↑ Cuando se cita algún autor como éstos es porque es el único que hizo la aclaración citada. Gifford, E. M., A. S. Foster 1989. Morphology and Evolution of Vascular Plants. Tercera edición. W. H. Freeman and Company. Nueva York. Capítulo 1. ISBN 0716719460
- ↑ a b Judd, WS; CS Campbell, EA Kellogg, PF Stevens, MJ Donoghue. 2007. Capítulo 7. An overview of green plant phylogeny. En: Plant Systematics, a phylogenetic approach. Third edition.
- ↑ a b PK Endress (2011) Evolutionary diversification of the flowers in angiosperms. Am. J. Bot. March 2011 vol. 98 no. 3 370-396 http://www.amjbot.org/content/98/3/370.full Archivado el 14 de diciembre de 2016 en Wayback Machine.
- ↑ Simpson, MG. 2005. Capítulo 11. Plant Embryology. En: Plant Systematics. Elsevier Academic Press.
- ↑ Simpson, MG. 2005. Capítulo 12. Palynology. En: Plant Systematics. Elsevier Academic Press.
- ↑ Simpson, MG. 2005. Capítulo 13. Plant Reproductive Biology. En: Plant Systematics. Elsevier Academic Press.
- ↑ González, A.M. y Arbo, M.M. «Sucesión foliar». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Archivado desde el original el 26 de agosto de 2012. Consultado el 16 de septiembre de 2012.
- ↑ González, A.M. y Arbo, M.M. «Porte». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 13 de septiembre de 2012.
- ↑ a b c Alicia Marticorena, Diego Alarcón, Lucía Abello y Cristian Atala. 2010. Plantas trepadoras, epífitas y parásitas nativas de Chile. Quinta guía de la serie de biodiversidad de CORMA. Ediciones Corporación Chilena de la Madera. Concepción, Chile. https://issuu.com/chilebosque/docs/marticorena_alarcon_abello_atala.2010.plantas_trep
- ↑ a b c González, A.M. y Arbo, M.M. «Plantas trepadoras». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 16 de septiembre de 2012.
- ↑ a b Mitraria coccinea, el Voqui, es una semitrepadora (arbusto apoyante). Enrique L. Ratera y Miguel O. Ratera. 1980. Plantas de la Flora Argentina empleadas en medicina popular. Editorial Hemisferio Sur S.A., Buenos Aires, Argentina.
- ↑ L Parodi (ed.) Enciclopedia Argentina de Agricultura y Jardinería, Tomo 1, las descripciones de plantas, utilizan el hábito de "arbusto apoyante", "arbusto trepador", "arbusto semitrepador", "arbusto semiapoyante".
- ↑ a b c d e Jackson, M. B. (ed.) 1986. New Root Formation in Plants and Cuttings. Serie Developments in plant and soil sciences nº 20. Martinus Nijhoff Publishers, a member of the Kluwer Academic Publishers Group. Da ordrecht / Boston / Lancaster. p.72. En p.74: los "banyan trees" tienen además "prop roots". p.77: Rhyzophora tiene "prop roots". Tabla en p.79: lista de especies con "prop roots". https://books.google.com.ar/books?id=gAv8CAAAQBAJ&pg=PA72
- ↑ a b c d e f g González, A.M. y Arbo, M.M. «Condiciones anormales de nutrición». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 16 de septiembre de 2012.
- ↑ http://www.biologia.edu.ar/botanica/tema20/21-1rizodermis.htm
- ↑ González, A.M. y Arbo, M.M. «Plantas epífitas». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 16 de septiembre de 2012.
- ↑ Ingrouille, Martin. 1992. Diversity and Evolution of land plants. Chapman & Hall.
- ↑ a b c d e f g h i González, A.M. y Arbo, M.M. «Adapataciones del cormo». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 16 de septiembre de 2012.
- ↑ a b c d e f González, A.M. y Arbo, M.M. «Raíz». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 15 de septiembre de 2012.
- ↑ a b «Raíces». Contexto Amazónico. Colombia: Universidad Nacional de Colombia, sede Amazonia. Archivado desde el original el 28 de septiembre de 2013. Consultado el 16 de septiembre de 2012.
- ↑ Crooks, Enos, Bank. 1997. [jxb.oxfordjournals.org/content/48/9/1703.full.pdf The function of buttress roots: a comparative study of the anchorage systems of buttressed (Aglaia and Nephelium ramboutan species) and non-buttressed (Mallotus wrayi) tropical trees.] J. Exp. Bot.
- ↑ a b González, A.M. y Arbo, M.M. «Plantas hidrófitas». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 16 de septiembre de 2012.
- ↑ a b Connie Barlow. 2000. "Why Pawpaw grows in patches". En: Connie Barlow. 2000. The Ghosts of Evolution. Nonsensical fruit, missing partners and other ecological anachronisms. First edition. Basic Books. A member of the Perseus Book Group.
- ↑ Watt, Michelle and John R. Evans (1999). «Proteoid roots. Physiology and developmengt». Plant Physiology 121: 317-323. doi:10.1104/pp.121.2.317. Consultado el 7 de noviembre de 2006.
- ↑ Grierson, P.F. and P. M. Attiwill (1989). «Chemical characteristics of the proteoid root mat of Banksia integrifolia L.». Australian Journal of Botany 37: 137-143. doi:10.1071/BT9890137.
- ↑ Milindasuta, B. 1975. Developmental anatomy of coralloid roots in Cycas. Amer. j. Bot. 62.
- ↑ Costa, J-L, P. Paulsrud, P. Lindblad. 1999. Cyanobiont diversity within coralloid roots of selected cycad species. FEMS Microbiology Ecology 28.
- ↑ Hallé, F., R. A. A. Oldeman, y P. B. Tomlinson. 1978. Trophic Trees and Forests: An Architectural Analysis. Springer, Berlin.
- ↑ González, A.M. y Arbo, M.M. «Sistemas de ramificación». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 13 de septiembre de 2012.
- ↑ González, A.M. y Arbo, M.M. «Tallo». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 13 de septiembre de 2012.
- ↑ Plant Ontology Consortium. Yemas vegetativas o reproductivas, las últimas florales o inflorescentes. No menciona las "yemas mixtas" de Simpson 2005 y Judd et al. 2007. http://www.plantontology.org/glossary
- ↑ a b c d e f g h i j k l m n González, A.M. y Arbo, M.M. «Yemas». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 13 de septiembre de 2012.
- ↑ Simpson (2005) relaciona la corteza (morfología) con un crecimiento secundario típico o anómalo, aquí se extiende la definición al crecimiento primario con posterior leñosidad, descripto en Valla (1979) Morfología de Plantas Superiores o en las familias expuestas en la flora mundial de Kubitzki (ed.); y se quitan los casos de crecimiento secundario sin posterior leñosidad, ya que los dos caracteres no están necesariamente acoplados.
- ↑ González, A.M. y Arbo, M.M. «Pecíolo». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Archivado desde el original el 31 de julio de 2013. Consultado el 16 de septiembre de 2012.
- ↑ Pemberton, R. W., y C. E. Turner. 1989. Ocurrence of predatory and fungivorous mites in leaf domatia. Am. J. Bot. 76: 105-112.
- ↑ Brouwer, Y. M., y H. T. Clifford. 1990. An annotated list of domatia-bearing species. Notes Jodrell Lab. 12: 1-50.
- ↑ Bentley, B. L. 1977. Extrafloral nectaries and protection by pugnacious bodyguards. Annu. Rev. Ecol. Syst. 8: 407-427.
- ↑ Hickey, L. J. 1973. Classification of the architecture of dicotyledoneous leaves. American Journal of Botany 60: 17-33.
- ↑ Hickey, L. J., y J. A. Wolfe. 1975. The bases of angiosperm phylogeny: vegetative morphology. Annals of the Missouri Botanical Garden 62: 538-589.
- ↑ González, A.M. y Arbo, M.M. «Partes constitutivas». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 10 de septiembre de 2012.
- ↑ Glosario del Angiosperm Phylogeny Website: http://www.mobot.org/mobot/research/apweb/top/glossarya_h.html
- ↑ "Capparaceae are woody, the inflorescence is racemose, the flowers have a long gynophore and often many stamens with long filaments, and the fruit is indehiscent." http://www.mobot.org/mobot/research/apweb/orders/brassicalesweb.htm#Capparoideae
- ↑ http://www.mobot.org/mobot/research/apweb/orders/brassicalesweb.htm#Cleomoideae
- ↑ a b c d e f Weberling, F. 1989. Morphology of flowers and inflorescences. (R. J. Pankhurst, trans.). Cambridge University Press, Cambridge. Preview en Google Books: http://books.google.com.ar/books?id=-ZlOAAAAIAAJ
- ↑ González, A.M. y Arbo, M.M. «Corola». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 10 de septiembre de 2012.
- ↑ Roth, I., & Lindorf, H. (1971). Desarrollo y anatomia del fruto y de la semilla de Theobroma cacao L. Acta Botánica Venezuélica, 6(1/4), 261-295. http://www.jstor.org/stable/41740588
- ↑ Morfología de la flor de Theobroma, incluyendo cogulla o capuchón petalar, en W Goitia, K Jaffé. Morfología comparativa de las flores de Theobroma cacao, T. bicolor, T. grandiflorum y Herrania sp. Acta Científica Venezolana 48. http://atta.labb.usb.ve/Klaus/art107.pdf
- ↑ a b c d Westerkamp C, Classen-Bockhoff R. 2007 Aug; Bilabiate flowers: the ultimate response to bees? Ann Bot. 100(2):361-74.
- ↑ Westerkamp C. 1997. Keel blossoms: bee flowers with adaptations against bees. Flora. 1997;192:125–132.
- ↑ a b c d González, A.M. y Arbo, M.M. «Androceo, cohesión y adnación». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 10 de septiembre de 2012.
- ↑ a b c González, A.M. «Androceo». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 10 de septiembre de 2012.
- ↑ Por ejemplo en Stuart Max Walters, James Cullen. 1986. The European Garden Flora: Pteridophyta, Gymnospermae, Angiospermae. p. 123: http://books.google.com.ar/books?id=1dd5M-ToXAcC&pg=PA123&lpg=PA123
- ↑ Gerald Carr en: http://www.biologie.uni-hamburg.de
- ↑ a b González, A.M. y Arbo, M.M. «Estilo y Estigma». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 10 de septiembre de 2012.
- ↑ González, A.M. y Arbo, M.M. «Inflorescencias simples». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 12 de septiembre de 2012.
- ↑ Faegri K, van der Pijl L. The principles of pollination ecology. 3rd edn. Oxford: Pergamon Press; 1979.
- ↑ a b c González, A.M. y Arbo, M.M. «Semilla». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 13 de septiembre de 2012.
- ↑ Duke J.A. 1969. On tropical tree seedlings. I. Seeds, seedlings, systems, and systematics. Ann.Missouri Bot.Gard. 56(2): 125-162.
- ↑ Tucker, A. O., M. J. Maciarello, y S. S. Tucker. 1991. A survey of color charts for biological descriptions. Taxon 40: 201-214.
- ↑ González, A.M. & Arbo, M.M. «Organización del cuerpo de la planta. Tema 2: Hoja. 2.2. Hoja de Dicotiledóneas». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 17 de setiembre de 2012.
- ↑ a b Systematics Association Committee for Descriptive Terminology. 1962. II. Terminology of Simple Symmetrical Plane Shapes (Chart 1). Taxon 11 (5): 145-156.
- ↑ a b c González, A.M. y Arbo, M.M. «Filotaxis». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Archivado desde el original el 29 de agosto de 2012. Consultado el 16 de septiembre de 2012.
- ↑ a b González, A.M. y Arbo, M.M. «Cáliz». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 12 de septiembre de 2012.
- ↑ González, A.M. y Arbo, M.M. «Plantas xerófitas». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 16 de septiembre de 2012.
- ↑ P. Font Quer, 1985. Diccionario de botánica. Editorial Labor, Barcelona.
- ↑ Font Quer, P., Diccionario de Botánica, Editorial Labor, Barcelona, 1985, p. 476.
- ↑ Handbook on seeds of dry-zone acacias, Anexo 6, Glosario, FAO, Departamento de Agricultura
<ref> definida en las <references> con nombre «UNNE gineceo ovario» no se utiliza en el texto anterior.Bibliografía
editarEl esqueleto de este texto tomado de
- Simpson, M. G. 2005 (segunda edición en 2010). "Plant Morphology". En: Plant Systematics. Elsevier Academic Press. Capítulo 9.
Completado con:
- Judd, Campbell, Kellogg, Stevens, Donoghue. 2007. Plant Systematics, a Phylogenetic approach. 3rd edition.
Simpson (2005) se basa en:
- Radford, A. E., W. C. Dickinson, J. R. Massey, y C. R. Bell. 1974. Vascular Plant Systematics. Harper & Row, Nueva York. (también recomendado por Judd et al. (2007), quienes agregan: "Incluye una lista de numerosos términos morfológicos y anatómicos, con ilustraciones.").
Y Simpson (2005) recomienda:
- Bell, A. D. 1991 (segunda edición en 2008). Plant Form, an Illustrated Guide to Flowering Plant Morphology. Oxford University Press, Oxford. La primera edición se puede desargar de https://archive.org/details/plantform00adri
Lecturas recomendadas por Judd et al. (2007), además de la ya comentada:
- Dilcher, D. 1974. Approaches to the identification of angiosperm leaf remains. Bot. Rev. 40:1-157. "Sumario muy detallado de los términos relacionados con las hojas de las angiospermas".
- Endress, P.K. 1994. Diversity and evolution of tropical flowers. Cambridge University Press, Cambridge. "Presentación detallada de la estructura y función de las flores tropicales".
- Hallé, F, RAA Oldeman y PB Tomlinson. 1978. Tropical trees and forests: An architectural analysis. Springer, Berlín.
- Harris, JG y MW Harris. 1994. Plant Identification Terminology: An illustrated glossary. Spring Lake Publishing, Spring Lake, UT.
- Hickey, LJ. 1973. Classification of the architecture of dicotyledonous leaves. Am. J. Bot. 60:17-33. "Sumario detallado relacionado con las hojas de las dicotiledóneas".
- Kiger, RW y DM Porter. 2001. Categorical glossary for the Flora of North America Project. Hunt Institute for Botanical Documentation, Carnegie Mellon University, Pittsburgh, PA.
- Weberling, F. 1989. Morphology of flowers and inflorescences. (RJ Pankhurst, trans.) Cambridge University Press, Cambridge. "Incluye una presentación detallada de la variación en la morfología floral y las inflorescencias, y términos asociados".
- Zomlefer, WB. 1994. Guide to flowering plant families. University of North Carolina Press, Chapel Hill. "Contiene un glosario hermosamente ilustrado de términos morfológicos comúnmente usados".
Bibliografía en español:
- Juan J. Valla (1ª ed. en 1979) Botánica. Morfología de Plantas Superiores. Hemisferio Sur.
Traducción de términos inglés-español en:
- Fernando Chiang, Mario Sousa S. y Mario Sousa P. Glosario Inglés-Español, Español-Inglés para Flora Mesoamericana Accedido 11/08/2013
 Datos: Q2911239
Datos: Q2911239 Multimedia: Botanical diagrams / Q2911239
Multimedia: Botanical diagrams / Q2911239