Giraffatitan brancai
Giraffatitan brancai es la única especie conocida del género extinto Giraffatitan de saurópodo macronario braquiosáurido que vivió a finales del periodo Jurásico, hace aproximadamente entre 150 a 145 millones de años, durante el Titoniense en lo que actualmente es África. Inicialmente se le consideró una especie africana de Brachiosaurus, Brachiosaurus brancai, pero estudios posteriores descartaron esta clasificación. Giraffatitan es uno de los mayores animales que caminó sobre la faz de la Tierra. Su morfología y fisiología ha podido ser ampliamente estudiada debido al hallazgo de un esqueleto casi completo y muy bien conservado correspondiente a este género.
| Giraffatitan brancai | ||
|---|---|---|
| Rango temporal: 150 Ma - 145 Ma Jurásico Superior | ||
 Esqueleto expuesto en el Museo de Historia Natural de Berlín, Alemania. | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| Superorden: | Dinosauria | |
| Orden: | Saurischia | |
| Suborden: | Sauropodomorpha | |
| Infraorden: | Sauropoda | |
| Familia: | Brachiosauridae | |
| Género: |
Giraffatitan Paul, 1988 | |
| Especie: |
G. brancai Janensch, 1914 | |
| Sinonimia | ||
| ||
Descripción
editarGiraffatitan brancai es un saurópodo de cuatro patas, herbívoro, de cuello largo, una cola relativamente corta y un cerebro y cráneo bastante pequeños. A diferencia de otros saurópodos, tenía una constitución similar a la de las actuales jirafas, con largas patas delanteras , en comparación a las traseras. Utilizaba su largo y robusto cuello para alimentarse de las copas de los árboles. Tenía unos dientes espatulados muy adecuados para su dieta. Su cráneo presenta bastantes agujeros, probablemente para reducir su peso, y una cresta característica. En cuanto a sus extremidades, el primer dedo de sus patas delanteras y los tres primeros de sus patas traseras tenían garras.[4]

Su contextura general es parecida a su pariente norteamericano Brachiosaurus altithorax, pero se puede apreciar un torso comparativamente más amplio en Brachiosaurus, y un cuello proporcionalmente más robusto y largo en Giraffatitan, además son apreciables numerosas diferencias anatómicas y proporcionales en el cráneo, las extremidades y la pelvis.[4][5][6]
Tamaño
editarDurante décadas, Giraffatitan fue considerado mundialmente como el mayor dinosaurio conocido. Pero posteriormente se han descubierto varios titanosaurios gigantes, como Argentinosaurus, que sobrepasaban el tamaño de Giraffatitan. Más recientemente, se ha descubierto otro saurópodo posiblemente braquiosáurido, Sauroposeidon, basado en un esqueleto incompleto, que parece muy razonable que supere a Giraffatitan.
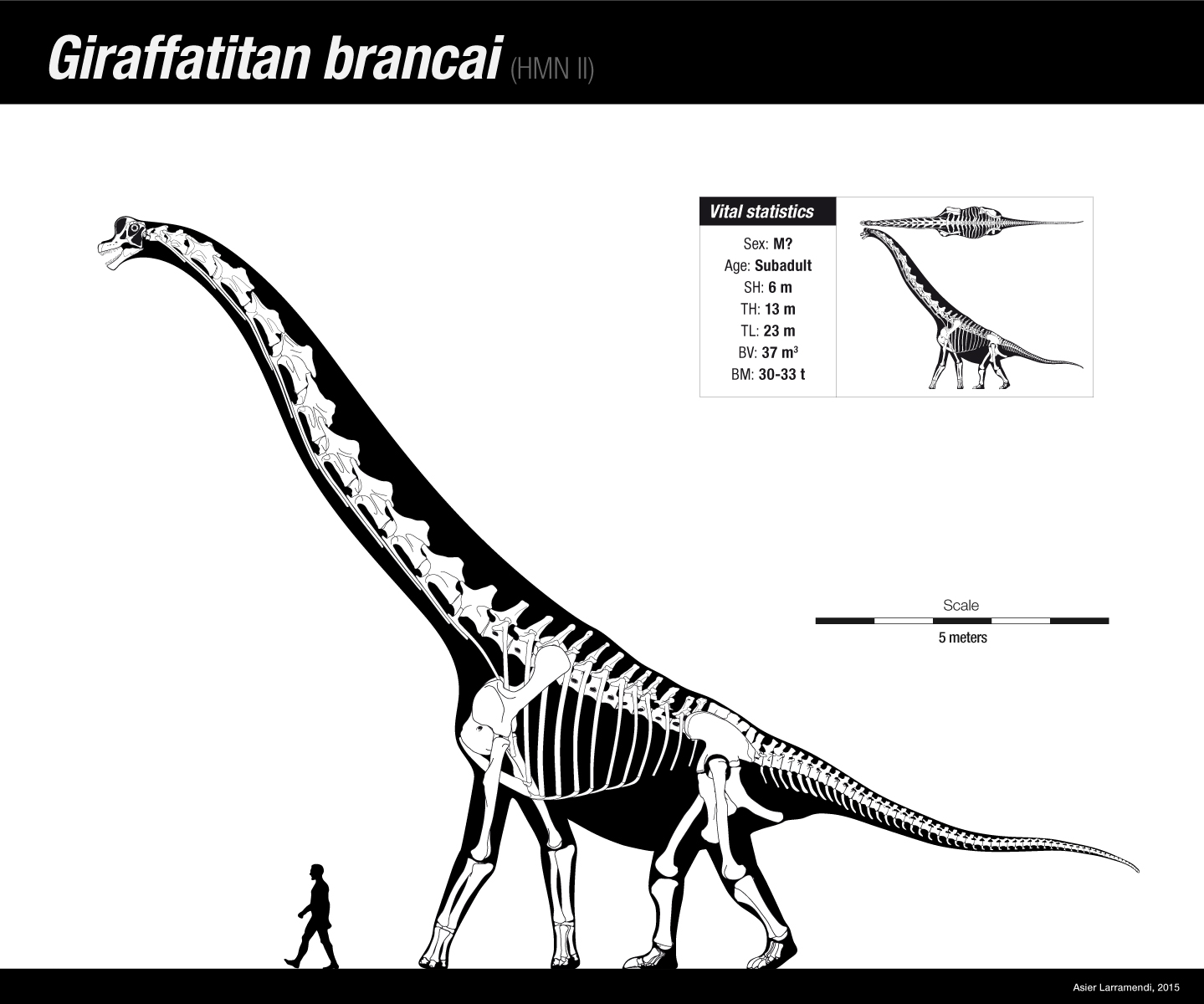
Basándose en el espécimen holotipo, HMN SII, conocido por un esqueleto casi completo que se expone en el Museo de Historia Natural de Berlín, Giraffatitan se estima que alcanzó entre 21,8 y 23 metros de longitud y probablemente medía 13 metros de altura hasta la cabeza.[7][8][9] Las estimaciones más recientes de su peso, basadas en modelos reconstruidos que toman en cuenta datos modernos de su osteología, como los sacos de aire y una finamente estimada masa muscular, calculan a este saurópodo en el rango de 23,2 a 39,5 toneladas.[3][4][9][8][10][11]

Se conoce un segundo espécimen fragmentario, HMN XV2, conocido básicamente por un hueso de la extremidad trasera, el peroné, que proporcionalmente es un 13% más grande que el correspondiente al holotipo. Este nuevo ejemplar supondría que Giraffatitan alcanzaría mayores tamaños,[4][12] rondando los 26 metros de largo y 15 de alto,[7] Gregory S. Paul estima su peso en 45 toneladas.[13]
Todas las estimaciones de tamaño para Giraffatitan se basan en el esqueleto montado en Berlín, en parte construido con huesos auténticos. Estos fueron tomados en gran parte del espécimen HMN SII,[4] un individuo subadulto entre 21,8 a 22,46 metros de longitud y unos 12 metros de altura.[8][14] La longitud a menudo mencionada de 22,46 metros es de Werner Janensch , el científico alemán que describió a Giraffatitan y fue el resultado de un simple error tipográfico el número correcto debería haber sido 22,16 metros. Las estimaciones de masa son más problemáticas e históricamente han variado fuertemente desde tan solo 15 toneladas hasta 78 toneladas. Estas estimaciones extremas ahora se consideran poco probables debido a metodologías defectuosas. También hay un gran número de estimaciones de este tipo, ya que el esqueleto demostró ser un tema irresistible para los investigadores que desean probar sus nuevos métodos de medición. Los primeros cálculos fueron hechos nuevamente por Janensch. En 1935, dio un volumen de treinta y dos metros cúbicos para el espécimen SII y de veinticinco metros cúbicos para el espécimen SI, un individuo más pequeño. No se sabe cómo llegó a estos números. En 1950, mencionó un peso de 40 toneladas para el esqueleto más grande.[14] En 1962, Edwin Harris Colbert midió un volumen de 86.953 m³. Suponiendo una densidad de 0,9, esto resultó en un peso de 78,258 kilogramos.[15] Colbert insertó un modelo de museo, vendido al público, en la arena y observó el volumen desplazado por ella. Gregory S. Paul en 1988 asumió que, en su opinión, un número demasiado realista había sido causado por el hecho de que tales modelos solían estar muy hinchados en comparación con la estructura real del animal.[3] En 1980, Dale Alan Russell publicó un peso mucho menor de 14,8 toneladas extrapolando el diámetro del húmero y el fémur.[16] En 1985, el mismo investigador llegó a 29 toneladas extrapolando de la circunferencia de estos huesos.[17] En 1985, Robert McNeill Alexander encontró un valor de 46,6 toneladas insertando un modelo de juguete del Museo Británico de Historia Natural en el agua.[18]
Las estimaciones más recientes basadas en modelos reconstruidos a partir de mediciones de volumen óseo, que tienen en cuenta los sistemas extensos de sacos aéreos reductores de peso presentes en los saurópodos, y la masa muscular estimada, están en el rango de 23 a 40 toneladas.[4][8] En 1988, Paul midió un volumen de 36,585 m³ insertando un modelo especialmente construido en el agua. Estimó un peso de 31,5 toneladas, asumiendo una baja densidad.[3] Entre 1994 y 1995, Jan Peczkis calculó un peso de cuarenta toneladas extrapolando a partir de la circunferencia del hueso del miembro.[19] En 1995, Hans-Christian Gunga utilizó un escáner láser.del esqueleto para construir un modelo virtual a partir de formas geométricas simples, encontrando un volumen de 74,42 m³ y concluyendo con un peso de 63 toneladas.[20] En 2008, Gunga revisó el volumen, utilizando formas más complejas, a 47,9 m³.[21] Donald Henderson en 2004 empleó un modelo de computadora que calculaba un volumen de 32,398 m³ y un peso de 25,789 kilogramos.[22] Los métodos más nuevos utilizan el espesor de la pared ósea.[23]
Fosas nasales
editar
Tradicionalmente, el distintivo cráneo de cresta alta se consideraba una característica del género Brachiosaurus, al que se refería originalmente Giraffatitan brancai.; Sin embargo, es posible que Brachiosaurus altithorax no muestre esta característica, ya que dentro del material tradicional de Brachiosaurus solo se conoce de especímenes tanzanianos ahora asignados a Giraffatitan.
La ubicación de las fosas nasales de Giraffatitan ha sido motivo de mucho debate con Witmer en 2001 que describe en Science la posición hipotética de las fosas nasales carnosas de Giraffatitan en tantas como cinco ubicaciones posibles. Comparando las narinas de los dinosaurios con las de los animales modernos, descubrió que todas las especies tienen sus aberturas nasales externas en el frente y que saurópodos como Giraffatitan no tenían fosas nasales sobre sus cabezas, sino cerca de sus hocicos.[24] También ha existido la hipótesis de varios saurópodos, como Giraffatitan, tuviesen una trompa. El hecho de que no hubo saurópodos de hocico estrecho, Giraffatitan incluido, tiende a desacreditar tal hipótesis. Se encuentra mayor evidencia de la ausencia de una trompa en el desgaste de los dientes de Giraffatitan, que muestra el tipo de desgaste que se produciría al morder y arrancar la materia vegetal en lugar de puramente moler, lo que sería el resultado de haber rasgado las hojas. y se ramifica con su trompa.[25]

La posición de las fosas nasales en la cabeza de Giraffatitan y en la mayoría de los saurópodos ha sido fuente controversia. Tradicionalmente, las grandes aberturas en la parte superior del cráneo han hecho pensar a los paleontólogos que era ahí donde se hallaban las fosas nasales. Pero estudios posteriores le han desacreditado fundamento a esta teoría de antaño, como en el artículo de Lawrence Witmer en 2001, que señala que la fosa nasal en posición rostral, en la parte delantera del hocico, es una regla prácticamente invariable en los animales amniotas. La comparación realizada entre las aberturas craneales de los dinosaurios y otros diápsidos modernos, como las tortugas, aves y cocodrilos, demostró que casi independientemente de la longitud, tamaño o posición relativa de los huesos nasales, el orificio nasal se encuentra en la posición más cercana posible del hocico, e incluso puede ser empujada más a esta posición con el recubrimiento cartilaginoso de la estructura ósea. Varios caracteres en el hocico de los saurópodos, como en el caso de Giraffatitan, sugiere que esta se encontraba óptimamente diseñada para la inserción de complejas estructuras carnosas y musculares, que hubiesen formado su fosa nasal, y posiblemente hasta una cámara de resonancia. Además la abundante presencia de finas y marcadas fisuras en el hocico, aluden a una abundante presencia de riego vascular, lo que aumenta la probabilidad que en realidad el orificio nasal estuviese en el hocico, en vez de al frente de los ojos.[26]
Descubrimiento e investigación
editar
Giraffatitan brancai fue nombrado y descrito por primera vez por el paleontólogo alemán Werner Janensch en 1914 como Brachiosaurus brancai, basado en varios especímenes recuperados entre 1909 y 1912 de la formación Tendaguru.[1] Se conoce a partir de cinco esqueletos parciales, incluidos tres cráneos y numerosos restos fragmentarios que incluyen material del cráneo, algunos huesos de las extremidades, vértebras y dientes. Vivió desde hace 145 hasta 150 millones de años, durante las edades de Kimmeridgiense a Titoniense del Jurásico Tardío.
Al describir el material del braquiosáurido de Tendaguru en 1914, Janensch enumeró una serie de diferencias y similitudes entre ellos y B. altithorax.[1] En otras tres publicaciones en 1929,[27] 1950[28] y 1961,[29] Janensch comparó las dos especies con más detalle, enumerando 13 características compartidas.[4] De estas, sin embargo, solo cuatro parecen ser válidas, mientras que seis pertenecen a grupos más inclusivos que Brachiosauridae, y el resto son difíciles de evaluar o se refieren a material que no es Brachiosaurus. En 1988, Gregory Paul publicó una nueva reconstrucción del esqueleto de "B." brancai, destacando una serie de diferencias en proporción entre este y B. altithorax. La principal es una diferencia en la forma en que varían las vértebras del tronco, son bastante uniformes en B. altithorax, pero varían ampliamente en el material africano. Paul creía que los elementos de las extremidades y cinturas de ambas especies eran muy similares, y por lo tanto sugirió separarlos no en el género, sino sólo en el nivel del subgénero.[3] Giraffatitan fue elevado a nivel de género por Olshevsky sin comentarios.[30] Un estudio detallado de todo el material, incluyendo los huesos de la extremidad y de la cintura, por Michael Taylor en 2009 encontró que hay diferencias significativas entre B. altithorax y el material de Tendaguru en todos los elementos conocidos de ambas especies. Taylor encontró 26 caracteres osteológicos distintos, una diferencia mayor que la existente entre, por ejemplo, Diplodocus y Barosaurus, y por lo tanto argumentó que el material africano debería ser colocado en su propio género, Giraffatitan, con la especie G. brancai. Una diferencia importante entre los dos géneros es la forma general del cuerpo, con Brachiosaurus teniendo un tronco 23% más largo y una cola 20 a 25% más larga y también más alta.[4]
Un famoso espécimen de Giraffatitan brancai montado en el Museo de Historia Natural de Berlín es uno de los esqueletos más grandes y más altos del mundo, como lo certifica el Libro Guinness de los Récords. A partir de 1909, Werner Janensch encontró muchos especímenes adicionales de G. brancai en Tanzania, África, incluidos algunos esqueletos casi completos, y los utilizó para crear el esqueleto montado compuesto que se ve hoy.
Canteras de Giraffatitan
editarEn 1906, el ingeniero de minas Bernhard Wilhelm Sattler, mientras viajaba, notó un enorme hueso que sobresalía del suelo en Tendagaru, la "colina empinada", cerca de Lindi, en lo que entonces era el África Oriental Alemana, hoy Tanzania. A principios de 1907, su superior Wilhelm Arning en Hannover recibió un informe sobre el hallazgo. Arning volvió a informar a la Kommission für die landeskundliche Erforschung der Schutzgebiete, una comisión en Berlín que analiza la investigación geográfica de los protectorados alemanes. El secretario de estado alemán de colonias, Berhard Dernburg, visitó entonces el África Oriental alemana acompañado por el industrial Heinrich Otto. Otto había invitado al paleontólogo Profesor Eberhard Fraas a unirse a él como asesor científico. En el verano de 1907, Fraas, que ya llevaba algunos meses viajando por la colonia, recibió una carta del Dr. Hans Meyer en Leipzig instándole a investigar el descubrimiento de Sattler. El 30 de agosto, Fraas llegó en vapor a la ciudad costera de Lindi. Una marcha de cinco días lo llevó a Tendagaru, donde pudo confirmar que los huesos eran auténticos y de dinosaurios. Pronto Sattler se unió a él con un equipo de mineros nativos que descubrieron dos grandes esqueletos de saurópodos que fueron transportados a Alemania. En última instancia, estos se convertirían en los holotipos de los géneros Tornieria y Janenschia.[31]
Fraas había observado que las capas de Tendaguru eran excepcionalmente ricas en fósiles. Después de su regreso a Alemania, trató de reunir suficiente dinero para una expedición importante. Logró atraer el interés del profesor Wilhelm von Branca, jefe de la Geologisch-Paläontologische Institut and Museum der Königliche Friedrich-Wilhelm Universität zu Berlin. Von Branca consideraba que era un orgullo nacional alemán que tal proyecto tuviera éxito. Participó el patólogo bien conectado David von Hansemann. Von Hansemann fundó un Comité de Tendaguru encabezado por Johann Albrecht, el duque de Mecklenburg. Pronto se puso de moda unirse a este comité que contaba con un gran número de prominentes industriales y científicos alemanes entre sus miembros. Muchos de sus amigos ricos donaron sumas considerables. Para dirigir la expedición, von Branca envió a uno de sus curadores, Werner Janensch y uno de sus asistentes, Edwin Hennig. Ambos llegaron a Dar es Salaam el 2 de abril de 1909.[31]
_(17544648673).jpg)
La expedición inicialmente empleó a unos 160 porteadores nativos, ya que las bestias de carga no podían usarse debido al peligro que representaba la mosca tse tse. Durante cuatro temporadas de campo, de 1909, 1910, 1911 y 1912, se abrieron alrededor de un centenar de canteras paleontológicas. Grandes cantidades de material fósil fueron enviadas a Alemania. Pronto se hizo evidente que aparte de Tornieria y Janenschia, otros saurópodos estaban presentes en las capas. Uno fue el Dicraeosaurus de tamaño mediano, un hallazgo relativamente común. Más rara era una forma gigantesca que superaba con creces a las otras en magnitud y que hoy se conoce como Giraffatitan. La primera cantera con material de Giraffatitan era el "Sitio D", ubicado aproximadamente a un kilómetro al noreste de la colina Tendaguru y se comenzó a excavar el 21 de junio de 1909. Contenía un esqueleto relativamente completo de un individuo de tamaño mediano, que carecía de las manos, el cuello, las vértebras y el cráneo. Incluía una serie articulada de veintinueve vértebras de la cola. Los otros huesos se encontraron en estrecha asociación en una superficie de 22 metros cuadrados. El "Sitio IX", ubicado a 1,4 kilómetros al noreste de la colina Tendaguru, se comenzó a excavar el 17 de agosto de 1909. Entre un conjunto de 150 huesos de dinosaurios desarticulados, también estaban presentes dos fémures de Giraffatitan. La próxima cantera de Giraffatitan fue el "Sitio N", a novecientos metros al este de la colina Tendaguru, excavada en septiembre de 1909. Contuvo un único esqueleto desarticulado que contenía una vértebra posterior, una vértebra de la cola, costillas y una escápula. una posible escápula, un húmero, dos isquias y varios huesos no identificables.[32]
La fuente más importante de fósiles de Giraffatitan sería el "Sitio S" a un kilómetro al suroeste de la colina. Las excavaciones comenzaron el 11 de octubre de 1909 y continuaron hasta 1912. En 1909 se desenterraron los elementos de las extremidades y la cintura. Durante 1910, una orilla cortada del río Kitukituki se profundizó gradualmente, eliminando una alta sobrecarga. Para evitar que las paredes de la cantera se derrumbaran, estaban cubiertas por un alto marco de madera. Ese año, primero se descubrieron varias costillas y luego parte de la columna vertebral. En octubre, cerca de algunas vértebras del cuello, se descubrieron un cráneo y mandíbulas inferiores. A partir del 5 de junio de 1912 se encontraron más vértebras en el cuello y tronco. Inicialmente se pensó que un solo esqueleto estaba siendo descubierto. Solo mucho después, Janensch se dio cuenta de que dos esqueletos habían estado presentes. El ejemplar SI estaba representado por un cráneo, seis vértebras del cuello y algunas vértebras de la espalda. El ejemplar SII era más grande, pero a pesar de su tamaño aún es un individuo subadulto. Incluía huesos del cráneo, una serie de once vértebras en el cuello y once en la espalda, costillas, escápula izquierda, ambos coracoides, ambas extremidades anteriores, los huesos púbicos y la extremidad posterior derecha. El sacro y la cola se habían perdido debido a una erosión relativamente reciente. El animal fue encontrado en posición vertical con las extremidades verticales, lo que se explica porque se habría quedado envuelto en lodo.[32]

A principios de octubre de 1909, se excavó el "Sitio ab", a 1,2 kilómetros al noreste de la colina. Entre los restos desarticulados de muchos saurópodos, también se recolectaron dos fémures de Giraffatitan. Un posible húmero gigantesco estaba demasiado dañado para ser salvado. El "sitio cc", a 2,9 kilómetros al noreste de la colina, contenía un esqueleto de Giraffatitan desarticulado que incluía vértebras del cuello, vértebras del tronco, costillas, escápula y húmero. En 1910, se abrió otra cantera de Giraffatitan, "Sitio Y" a 3.1 kilómetros al norte de la colina de Tendaguru. Contenía el esqueleto de un individuo de tamaño mediano que incluía una base de cerebro, una serie de ocho vértebras del cuello, una vértebra del tronco, costillas, ambas escápulas, un coracoides, un húmero izquierdo y un peroné izquierdo.[32]
Las canteras mencionadas anteriormente representan solo los sitios más importantes donde se encontraron huesos de Giraffatitan. En docenas de otras ubicaciones de Tendagaru se encontraron hallazgos de huesos grandes de saurópodos que fueron referidos al taxón en las publicaciones de Janensch, pero de los cuales no se conservan las notas de campo, por lo que se desconocen las circunstancias precisas de los descubrimientos. En parte, esto refleja una falta de documentación sistemática por parte de la expedición. Muchos bombardeos aliados destruyeron muchos documentos en 1943. También se perdió parte de los fósiles. Sin embargo, la mayor parte del esqueleto es conocido.[4]
Clasificación
editar
Giraffatitan brancai se clasificó inicialmente como una especie de Brachiosaurus, según Janensch, en 1914. Se le conoce por cinco esqueletos parciales, que incluyen al menos tres cráneos, recogidos cerca de Lindi, Tanzania, cerca de 1900. Vivió entre 145 y 150 millones de años atrás. En 1988, Gregory S. Paul notó que la forma africana, en el cual se basaban las representaciones más populares de Brachiosaurus, mostraba diferencias significativas con la forma americana, principalmente en sus vértebras y en su constitución más grácil. Paul usó estas diferencias para crear el subgénero que llamó Brachiosaurus (Giraffatitan) brancai. En 1991, George Olshevsky aseveró que las diferencias eran suficientes para crear el género Giraffatitan.[33]

La descripción revisada de un cráneo de Brachiosaurus norteamericano, en 1998, aumentó las diferencias. Este cráneo, encontrado una centuria antes, es el cráneo que Marsh empleó en sus reconstrucciones de Apatosaurus, se identifica como "Brachiosaurus sp." y podría pertenecer a B. altithorax. El cráneo está más cerca de Camarasaurus en algunas características, como la forma de los dientes frontales y más alargado y menos ahuecado en la parte superior que el distintivo cráneo de Giraffatitan, de hocico corto y de cresta alta . .[34]
La clasificación de Giraffatitan como género separado no fue muy popular entre los científicos al principio, al no basarse en comparaciones rigurosas. Sin embargo, Michael Taylor publicó una comparación detallada en 2009. Taylor mostró que "Brachiosaurus" brancai difería de B. altithorax en casi cada uno de los huesos comparados, y en todos los aspectos, validando Giraffatitan como un género separado.[4] Taylor encontró evidencia de una relación hermana entre Giraffatitan y Brachiosaurus, aunque su análisis omitió otros braquiosáuridos.[4] Un estudio más reciente sobre los saurópodos titanosauriformes de D'Emic de 2012 coloca a Giraffatitan como un clado hermano Brachiosaurus y un tritomio que contiene de Abydosaurus, Cedarosaurus y Venenosaurus.[35]
Filogenia
editarCladograma de la familia Brachiosauridae, basado en el análisis filogenético de Michae D'Emic en 2012. Mostrando la posición sistemática de Giraffatitan.[35]
| Brachiosauridae |
| |||||||||||||||||||||||||||
Paleobiología
editarCerebro
editarComo otros saurópodos, Giraffatitan tenía un cerebro relativamente pequeño, incluso cuando se considera su enorme tamaño corporal. Su bóveda craneal, espacio ocupado esencialmente por el cerebro, tenía un volumen de solo de 300 cm³, mientras que en los humanos ronda los 1350 cm³ y en Tyrannosaurus rex los 343 cm³.[36][37] Un estudio de 2009 calculaba su cociente cerebro/cuerpo, un cálculo aproximado de la inteligencia posible, entre un 0,62 a 0,79, dependiendo de la estimación del tamaño utilizado. Giraffatitan es también similar a otros saurópodos en tener una ampliación del canal espinal sobre las caderas, lo que algunas de las fuentes antiguas refirieron engañosamente como "segundo cerebro".[38]
Metabolismo
editarSi Giraffatitan hubiese sido endotérmico, de sangre caliente, habría tardado 10 años en alcanzar su madurez, pero si hubiese sido poiquilotermo, de sangre fría, habría necesitado más de 100 años.[39] Como animal de sangre caliente, las demandas energéticas diarias de un braquiosaurio habrían sido enormes, necesitando probablemente comer más de 182 kilogramos de alimento por día. En cambio, si el Giraffatitan era completamente de sangre fría o de una pasiva gigantotermia, mantenerse calientes debido a su enorme tamaño, habría necesitado mucho menos alimento para cubrir sus necesidades energéticas diarias. Algunos científicos han propuesto que los dinosaurios grandes como el Giraffatitan se mantenían calientes por su enorme tamaño.[40]
Medio ambiente y comportamiento
editar
Junto con la antigua teoría de que los orificios nasales de Ssaurópodos como Giraffatitan se encontraban por encima de los ojos, se pensaba que estas funcionaban como una especie de esnórquel natural para respirar bajo el agua, ya que se consideraba que estos animales tenían que permanecer bajo el agua para sostener su enorme peso. Pero estudios posteriores han demostrado que la presión del agua en realidad habría impedido que el animal respire adecuadamente mientras estaba sumergido, y que sus pies eran demasiado estrechos para su uso eficiente en el fangoso suelo acuático. Además también se ha demostrado, mediante estudios de simulación biomecánica por ordenador, que animales terrestres cuadrúpedos mucho mayores que Giraffatitan, como Argentinosaurus, sí eran capaces de caminar eficientemente en suelo terrestre y que sus articulaciones soportasen su gran masa corporal.[41] Giraffatitan fue uno de los más grandes dinosaurios en el Jurásico, viviendo en praderas llenas de helechos, bennetites y equisetos, moviéndose y alimentándose entre los vastos bosques de coníferas y cícadas, helechos con semilla y ginkgos.
Paleoecología
editarGiraffatitan vivió en lo que hoy es Tanzania en la Formación Jurásica tardía de Tendaguru.[42] Desde 2012, el límite entre Kimmeridgiano y Titoniano está fechado en 152.1 millones de años.[43] El ecosistema de Tendaguru consistía principalmente en tres tipos de entornos, ambientes marinos someros, como lagunas, zona intermareal y ambientes costeros bajos y en ambientes interiores con vegetación. El ambiente marino existía por encima de la base de la ola y detrás de las barreras de siliciclasticos y ooides. Parecía haber tenido pocos cambios en los niveles de salinidad y mareas y tormentas experimentadas. Los ambientes costeros consistían en lagos, estanques y pozas costeras salobres. Estos ambientes tenían poca vegetación y probablemente fueron visitados por dinosaurios herbívoros, principalmente durante las sequías. Las tierras con buena vegetación estaban dominadas por coníferas. En general, el clima del Jurásico tardío fue subtropical a tropical con lluvias estacionales y períodos secos pronunciados. Durante el Cretácico temprano , el Tendaguru se volvió más húmedo.[44] Las camas Tendaguru son similares a la Formación Morrison de América del Norte, excepto en sus espacios marinos.[45]
Giraffatitan habría coexistido con compañeros saurópodos como Dicraeosaurus hansemanni y D. sattleri, Janenschia africana, Tendaguria tanzaniensis y Tornieria africanus. Ornitisquios como Dysalotosaurus lettowvorbecki y Kentrosaurus aethiopicus. Los terópodos "Allosaurus" tendagurensis, Ceratosaurus roechlingi, "Ceratosaurus" ingens, Elaphrosaurus bambergi, Veterupristisaurus milneri y Ostafrikasaurus crassiserratus y el pterosaurio Tendaguripterus recki.[46][47][48][49] Otros organismos que habitaron el Tendaguru incluían corales, equinodermos, cefalópodos, bivalvos, gastrópodos, decápodos, tiburones, peces neopterygios, cocodrilos y pequeños mamíferos como Brancatherulum tendaguerense.[50]
En la cultura popular
editarGiraffatitan es uno de los dinosaurios más conocidos entre los paleontólogos y el público en general, aunque con el antiguo nombre Brachiosaurus brancai. Un famoso espécimen de Giraffatitan brancai montado en el Museo de Historia Natural de Berlín es uno de los mayores, y de hecho el más alto, de los esqueletos que se han montado en el mundo, como acredita el libro Guinness de los récords, con una altura de 13,27 metros y 22,5 metros de largo. El esqueleto está compuesto a partir de huesos de distintos ejemplares proporcionados por las campañas de Werner Janensch en Tanzania.
Referencias
editar- ↑ a b c Janensch, W. (1914). "Übersicht über der Wirbeltierfauna der Tendaguru-Schichten nebst einer kurzen Charakterisierung der neu aufgeführten Arten von Sauropoden." Archiv für Biontologie, 3 (1): 81–110.
- ↑ Kingham, R. F. (1962). «Studies of the sauropod dinosaur Astrodon Leidy». Washington Junior Academy of Science Proceedings, 1: 38-44 Resumen de la taxonomía en Paleobiology Database
- ↑ a b c d e Paul, G.S. (1988). «The brachiosaur giants of the Morrison and Tendaguru with a description of a new subgenus, Giraffatitan, and a comparison of the world's largest dinosaurs». Hunteria 2 (3): 1-14.
- ↑ a b c d e f g h i j k Taylor, M.P. (2009). "A Re-evaluation of Brachiosaurus altithorax Riggs 1903 (Dinosauria, Sauropod) and its generic separation from Giraffatitan brancai (Janensh 1914)." Journal of Vertebrate Paleontology, 29(3): 787-806.
- ↑ Hartman, Scott (2014). «Sauropods and Kin: Brachiosaurus altithorax». Scott Hartman's Skeletal drawing.com. Consultado el 7 de julio de 2016.
- ↑ Hartman, Scott (2014). «Sauropods and Kin: Giraffatitan brancai». Scott Hartman's Skeletal drawing.com. Consultado el 7 de julio de 2016.
- ↑ a b Taylor, Mike (2 de junio de 2014). «How big was Brachiosaurus altithorax? I mean, really?». Sauropod Vertebra Picture of the Week. Consultado el 7 de julio de 2016.
- ↑ a b c d Mazzetta, G.V. (2004). «Giants and Bizarres: Body Size of Some Southern South American Cretaceous Dinosaurs.». Historical Biology 16: 1-13. doi:10.1080/08912960410001715132.
- ↑ a b Larramendi, Asier (2015). «Giraffatitan brancai». Consultado el 7 de julio de 2016.
- ↑ Christiansen, P. (1997). "Feeding mechanisms of the sauropod dinosaurs Brachiosaurus, Camarasaurus, Diplodocus and Dicraeosaurus." Historical Biology, 14(3): 137-152.
- ↑ Campione, Nicolás E.; Evans, David C. (10 de julio de 2012). «A universal scaling relationship between body mass and proximal limb bone dimensions in quadrupedal terrestrial tetrapods». BioMed Central Biology 10 (1): 1. doi:10.1186/1741-7007-10-60. Consultado el 7 de julio de 2016.
- ↑ Hartman, Scott (22 de julio de 2014). «Smackdown: Supersaurus vs. Giraffatitan and Diplodocus». Scott Hartman's Skeletal drawing.com. Consultado el 7 de julio de 2016.
- ↑ Paul, Gregory S. (Marzo de 2010). «Gregory S. Paul's Dinosaur Mass Table». Consultado el 7 de julio de 2016.
- ↑ a b Janensch, W. (1950). The Skeleton Reconstruction of Brachiosaurus brancai. pp. 97-103.
- ↑ Colbert, E (1962). «The weights of dinosaurs». American Museum Novitates 2076: 1-16.
- ↑ Russell, D.; Béland, P.; McIntosh, J.S. (1980). «Paleoecology of the dinosaurs of Tendaguru (Tanzania)». Mémoires de la Societé géologique de la France 59: 169-175.
- ↑ Anderson, J.F.; Hall-Martin, A.; Russell, D.A. (1985). «Long-bone circumference and weight in mammals, birds and dinosaurs». Journal of Zoology 207 (1): 53-61. doi:10.1111/j.1469-7998.1985.tb04915.x.
- ↑ Alexander, R. McN. (1985). «Mechanics of posture and gait of some large dinosaurs». Zoological Journal of the Linnean Society 83 (1): 1-25. doi:10.1111/j.1096-3642.1985.tb00871.x.
- ↑ Peczkis, J. (1995). «Implications of body-mass estimates for dinosaurs». Journal of Vertebrate Paleontology 14 (4): 520-533.
- ↑ Gunga, H.-C.; Kirsch, K.A.; Baartz, F.; Röcker, L.; Heinrich, W.-D.; Lisowski, W.; Wiedemann, A.; Albertz, J. (1995). «New data on the dimensions of Brachiosaurus brancai and their physiological implications». Naturwissenschaften 82 (4): 190-192.
- ↑ Gunga, H.-C.; Suthau, T.; Bellmann, A.; Stoinski, S.; Friedrich, A.; Trippel, T.; Kirsch, K.; Hellwich, O. (2008). «A new body mass estimation of Brachiosaurus brancai Janensch, 1914 mounted and exhibited at the Museum of Natural History (Berlin, Germany)». Fossil Record 11 (1): 33-38. doi:10.1002/mmng.200700011.
- ↑ Henderson, D.M. (2004). «Tipsy Punters: Sauropod Dinosaur Pneumaticity, Buoyancy and Aquatic Habits». Proceedings: Biological Sciences 271 (Supplement 4): 180-183.
- ↑ Benson, R. B. J.; Campione, N. S. E.; Carrano, M. T.; Mannion, P. D.; Sullivan, C.; Upchurch, P.; Evans, D. C. (2014). «Rates of Dinosaur Body Mass Evolution Indicate 170 Million Years of Sustained Ecological Innovation on the Avian Stem Lineage». PLoS Biology 12 (5): e1001853. PMC 4011683. PMID 24802911. doi:10.1371/journal.pbio.1001853.
- ↑ Witmer, L.M. (2001). «Nostril position in dinosaurs and other vertebrates and its significance for nasal function». Science 293 (5531): 850-853. PMID 11486085. doi:10.1126/science.1062681. Archivado desde el original el 6 de septiembre de 2013.
- ↑ Naish, D. (20 de marzo de 2009). «Junk in the trunk: why sauropod dinosaurs did not possess trunks». ScienceBlogs. Archivado desde el original el 13 de enero de 2012.
- ↑ Witmer, Lawrence (2001). «Nostril position in dinosaurs and other vertebrates and its significance for nasal function». Science 293 (5531): 850-853. Archivado desde el original el 16 de septiembre de 2016. Consultado el 7 de julio de 2016.
- ↑ Janensch, W. (1929). "Material und Formengehalt der Sauropoden in der Ausbeute der Tendaguru-Expedition." Palaeontographica (Suppl. 7) 2:1–34.
- ↑ Janensch, W. (1950). "Die Wirbelsäule von Brachiosaurus brancai." Palaeontographica (Suppl. 7) 3:27–93.
- ↑ Janensch, W. (1961). "Die Gliedmaßen und Gliedmaßengürtel der Sauropoden der Tendaguru-Schichten." Palaeontographica (Suppl. 7) 3:177–235.
- ↑ Olshevsky, G (1991). «A revision of the parainfraclass Archosauria Cope, 1869, excluding the advanced Crocodylia». Mesozoic Meanderings 2: 1-196.
- ↑ a b Maier, Gerhard. 2003. African dinosaurs unearthed: the Tendaguru expeditions. Life of the Past Series (ed. J. Farlow). Indiana University Press, Bloomington, Indiana
- ↑ a b c Wolf-Dieter Heinrich, 1999, "The Taphonomy of Dinosaurs from the Upper Jurassic of Tendaguru (Tanzania) Based on Field Sketches of the German Tendaguru Expedition (1909-1913)", Mitteilungen aus dem Museum für Naturkunde in Berlin. Geowissenschaftliche Reihe 2: 25-61
- ↑ Glut, D.F. (1997). «Brachiosaurus». Dinosaurs: The Encyclopedia. McFarland & Company. p. 218. ISBN 0-89950-917-7.
- ↑ Carpenter, K. and Tidwell, V. (1998). "Preliminary description of a Brachiosaurus skull from Felch Quarry 1, Garden Park, Colorado." Pp. 69–84 in: Carpenter, K., Chure, D. and Kirkland, J. (eds.), The Upper Jurassic Morrison Formation: An Interdisciplinary Study. Modern Geology, 23(1-4).
- ↑ a b D'Emic, M. D. (2012). «The early evolution of titanosauriform sauropod dinosaurs». Zoological Journal of the Linnean Society 166 (3): 624-671. doi:10.1111/j.1096-3642.2012.00853.x.
- ↑ Ross, Callum; Henneberg, Maciej (1995). «Basicranial flexion, relative brain size, and facial kyphosis in Homo sapiens and some fossil hominids». American Journal of Physical Anthropology 98 (4): 575-593. Consultado el 7 de julio de 2016.
- ↑ Carabajal, Ariana; Canale, Juan (Septiembre de 2010). «Cranial endocast of the carcharodontosaurid theropod Giganotosaurus carolinii CORIA & SALGADO, 1995». Neues Jahrbuch für Geologie und Paläontologie (E. Schweizerbart'sche Verlagsbuchhandlung) 258 (2): 249-256. doi:10.1127/0077-7749/2010/0104. Consultado el 7 de julio de 2015.
- ↑ Knoll, F. y Schwarz-Wings, D.(2009). «Palaeoneuroanatomy of Brachiosaurus». Annales de Paléontologie, 95(3): 165-175 doi: 10.1016/j.annpal.2009.06.001.
- ↑ Case, T.J. (1978). «Speculations on the Growth Rate and Reproduction of Some Dinosaurs». Paleobiology 4 (3): 323.
- ↑ Bailey, J.B. (1997). "Neural spine elongation in dinosaurs: Sailbacks or buffalo-backs?" Journal of Paleontology, 71(6): 1124-1146.
- ↑ Sellers, William; Margetts, Lee; Coria, Rodolfo; Manning, Phillip (30 de octubre de 2013). «March of the Titans: The Locomotor Capabilities of Sauropod Dinosaurs». PLoS ONE 8 (10). PMID 24348896. doi:10.1371/journal.pone.0078733. Consultado el 7 de julio de 2016.
- ↑ Robert Bussert, Wolf-Dieter Heinrich and Martin Aberhan, 2009, "The Tendaguru Formation (Late Jurassic to Early Cretaceous, southern Tanzania): definition, palaeoenvironments, and sequence stratigraphy", Fossil Record 12(2) 2009: 141–174
- ↑ Gradstein, F.M.; Ogg, J.G.; Schmitz, M.D. & Ogg, G.M., 2012, A Geologic Time Scale 2012, Elsevier
- ↑ Aberhan, Martin; Bussert, Robert; Heinrich, Wolf-Dieter; Schrank, Eckhart; Schultka, Stephan; Sames, Benjamin; Kriwet, Jürgen; Kapilima, Saidi (2002). «Palaeoecology and depositional environments of the Tendaguru Beds (Late Jurassic to Early Cretaceous, Tanzania)». Fossil Record 5 (1): 19-44. doi:10.1002/mmng.20020050103.
- ↑ Mateus, Octávio (2006). «Late Jurassic dinosaurs from the Morrison Formation (USA), the Lourinhā and Alcobaça formations (Portugal), and the Tendaguru Beds (Tanzania): a comparison». En Foster, J.R.; Lucas, S.G., eds. Paleontology and Geology of the Upper Morrison Formation. New Mexico Museum of Natural History and Science (bulletin 36). pp. 223-232. ISSN 1524-4156.
- ↑ Weishampel, David B; et al. (2004). "Dinosaur distribution (Late Jurassic, Africa)." In: Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.): The Dinosauria, 2nd, Berkeley: University of California Press. p. 552. ISBN 0-520-24209-2.
- ↑ Barrett, P.M., Butler, R.J., Edwards, N.P., & Milner, A.R. Pterosaur distribution in time and space: an atlas. p61–107. in Flugsaurier: Pterosaur papers in honour of Peter Wellnhofer. 2008. Hone, D.W.E., and Buffetaut, E. (eds). Zitteliana B, 28. 264pp.[1]
- ↑ Rauhut, Oliver W. M. (2011). «Theropod dinosaurs from the Late Jurassic of Tendaguru (Tanzania)». Special Papers in Palaeontology 86: 195-239. doi:10.1111/j.1475-4983.2011.01084.x.
- ↑ Buffetaut, Eric (2012). «An early spinosaurid dinosaur from the Late Jurassic of Tendaguru (Tanzania) and the evolution of the spinosaurid dentition». Oryctos 10: 1-8.
- ↑ Heinrich, Wolf-Dieter (2001). «The German‐Tanzanian Tendaguru Expedition 2000». Fossil Record 4 (1): 223-237. doi:10.1002/mmng.20010040113.
{kind=link}
 Datos: Q124481950
Datos: Q124481950